Статья: Что и как закодировано в мРНК
Все приведенные примеры нарушения общих правил кодирования так или иначе связаны с существованием определенного контекста в мРНК. Этот контекст или перекодирующие сигналы иногда называют вторым генетическим кодом.
МНОГИЕ мРНК СОДЕРЖАТ СИГНАЛЫ ПОЛИАДЕНИЛИРОВАНИЯ
Около половины мРНК эукариот полиаденилируются на 3'-конце во время процессинга в клеточном ядре. Сигналом ядерного полиаденилирования 3'-конца служит последовательность AAUAAA, расположенная за 10-20 нуклеотидов от этого конца. В некоторых случаях, например в зрелых ооцитах амфибий или сухих, неактивных в белковом синтезе зародышах пшеницы, мРНК не полиаденилированы. Активация белкового синтеза в развитии сопровождается полиаденилированием этих мРНК в цитоплазме. Для цитоплазматического полиаденилирования мРНК должны содержать в 3'-нетранслируемой области наряду с элементом ядерного полиаденилирования дополнительную последовательность - элемент цитоплазматического полиаденилирования. Такой последовательностью у амфибий является (U)6AUAAAG. Поли(А)-хвост на мРНК узнается особым поли(А)-связывающим белком, который участвует в инициации трансляции мРНК по кэп-зависимому механизму.
СПЕЦИФИЧЕСКИЕ ПОСЛЕДОВАТЕЛЬНОСТИ В мРНК ОТВЕЧАЮТ ЗА РЕГУЛЯЦИЮ ЕЕ МАТРИЧНОЙ АКТИВНОСТИ В БЕЛКОВОМ СИНТЕЗЕ
Матричная активность различных мРНК сильно различается. Очень активными (сильными) матрицами являются фаговые и вирусные РНК, а также клеточные мРНК для мажорных белков клетки, таких, например, как глобины. Наоборот, матрицы для белков, присутствующих в клетке в малых количествах, как правило, являются очень слабыми. Сила матрицы чаще всего определяется эффективностью процесса ее инициации. У эукариот помимо контекста нуклеотидов в районе инициирующего триплета (см. выше) эффективность матрицы сильно падает при наличии в 5'-нетранслируемой области развитой вторичной структуры (образовании двуспиральных участков, таких, как шпильки), а также, по-видимому, с увеличением длины 5'-НТО выше определенного предела. Активность эукариотических мРНК в трансляции катастрофически уменьшается при их декэпировании или введении в 5'-НТО кодонов AUG, в контексте, неоптимальном для инициации. Прокариотические мРНК плохо транслируются, если инициирующие кодоны находятся в двойных спиралях.
Избирательное влияние на активность мРНК в трансляции оказывают специфические регуляторные белки или специальные регуляторные РНК. Эти белки или РНК проявляют свое действие, связываясь со специфическими последовательностями или структурами в мРНК, которые называются регуляторными элементами. В большинстве случаев регуляторные элементы располагаются в 5'-НТО вблизи инициирующего кодона. Однако в некоторых случаях регулярные элементы могут быть на значительном расстоянии от инициирующего кодона, в том числе в 3'-НТО. Связываясь с мРНК вблизи инициирующего кодона, регуляторные белки могут создавать препятствия для компонентов белоксинтезирующего аппарата (мешать связыванию с мРНК или ее сканированию). При связывании с мРНК на большом расстоянии от места инициации регуляторные белки могут влиять на процесс инициации путем изменения общей пространственной структуры мРНК, изменяя таким образом доступность инициирующего кодона или 5'-конца мРНК для белоксинтезирующего аппарата.
Регуляторными белками могут быть специальные белки клетки, выполняющие только эту функцию в организме, а также белки, имеющие в организме другие функции и работающие в качестве регуляторных белков "по совместительству". Довольно часто в качестве белков, регулирующих активность определенных мРНК, могут выступать сами продукты их трансляции (авторегуляция). Обратимся теперь к некоторым конкретным примерам.
Трансляция мРНК треонил-тРНК-синтетазы у бактерий находится под контролем ее продукта (авторегуляция). Авторегуляция достигается за счет специфического сродства треонил-тРНК-синтетазы к своей матрице. Связывание синтетазы с мРНК происходит в ее 5'-НТО со специфической последовательностью (регуляторным элементом), которая складывается во вторичную структуру, напоминающую отдельные элементы вторичной структуры треониловой тРНК. При ограниченном количестве фермента в клетке он ассоциирует главным образом со своим субстратом - треонил-тРНК. В этих условиях мРНК треонил-тРНК-синтетазы может активно транслироваться. При увеличении количества этого фермента избыточный фермент взаимодействует с регуляторным элементом в 5'-НТО своей мРНК. Это приводит к переходу мРНК в такую конформацию, в которой инициирующий триплет оказывается вовлеченным в двуспиральную структуру. В результате инициация синтеза треонил-тРНК-синтетазы прекращается.
Яркий пример регуляции трансляции мРНК в эукариотических клетках получен при изучении механизмов поддержания в клетках уровня свободного железа. Железо входит в состав активных центров очень многих белков, таких, например, как гемоглобин , миоглобин, цитохромы , однако ионы свободного железа токсичны для клетки и поэтому связываются и переводятся в нетоксичную форму белком ферритином. Синтез ферритина в клетке, в свою очередь, зависит от уровня свободного железа: в присутствии железа ферритин синтезируется, в то время как при его недостатке трансляция мРНК ферритина останавливается на стадии инициации. Выяснилось, что регуляция синтеза ферритина целиком зависит от специфической последовательности длиной 26 нуклеотидов, образующей шпилечную структуру в 5'-НТО мРНК ферритина (рис. 4, а). Этот регуляторный элемент при отсутствии железа связывается со специфическим белком, который препятствует сканированию 5'-НТО рибосомами и, таким образом, подавляет трансляцию мРНК на стадии ее инициации. Этот белок- репрессор имеет сродство к ионам железа и при связывании с ними перестает связываться с ферритиновой мРНК. После диссоциации белка мРНК становится активной в синтезе ферритина.
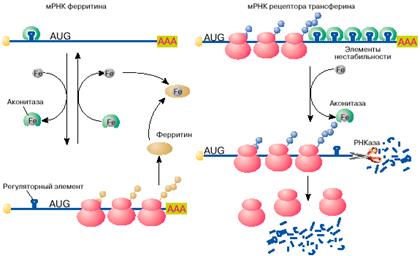
Рис. 4. Регуляция железом трансляции мРНК ферритина и стабильности мРНК рецептора трансферина
Вновь синтезированный ферритин отнимает железо у репрессора. Утратив железо, репрессор опять приобретает сродство к регуляторному элементу ферритиновой мРНК, связывается с ним и останавливает синтез ферритина. Большим сюрпризом явилось то, что белок-репрессор оказался хорошо известным ферментом цикла Кребса - аконитазой.
ОСОБЫЕ СТРУКТУРНЫЕ ЭЛЕМЕНТЫ мРНК ОПРЕДЕЛЯЮТ ВРЕМЯ ЖИЗНИ мРНК В КЛЕТКЕ
Скорость синтеза белка зависит от количества соответствующей мРНК в клетке. Содержание мРНК, в свою очередь, определяется скоростью ее синтеза и распада. Механизмы регуляции транскрипции мРНК начали исследовать еще в начале 60-х годов, механизмы регуляции процесса ее деградации - значительно позже, всего 5-10 лет назад.
Время жизни разных мРНК в клетках эукариот варьирует в очень широких пределах. Период их полураспада составляет от десятков минут до десятка суток. Наиболее стабильными являются мРНК, на которых синтезируются белки, обеспечивающие функционирование организма как целого (например, мРНК сывороточного альбумина ). Некоторые мРНК распадаются на определенных стадиях клеточного цикла или определенных этапах процесса клеточной дифференцировки. Время жизни мРНК в клетке, равно как и момент их деградации, запрограммировано специфическими последовательностями или структурами в мРНК, часто в их 3'-нетранслируемой области. Эти последовательности (структуры) обычно узнаются специфическими белками. Их связывание с соответствующими белками обычно приводит к стабилизации мРНК.
Например, за деградацию гистоновых мРНК на определенной стадии клеточного цикла отвечает особая шпилечная структура в 3'-НТО - элемент нестабильности мРНК. Введение такой структуры в 3'-НТО очень стабильной глобиновой мРНК приводит к ее деградации одновременно с гистоновыми мРНК. Для того чтобы эта структура выполняла свою функцию элемента нестабильности, она должна находиться на определенном, достаточно близком расстоянии от терминирующего кодона. Дестабилизирующим действием в мРНК онкобелка c- myc обладает AU-богатая последовательность длиной 60-80 нуклеотидов в 3'-НТО этой мРНК. В этой последовательности был выявлен многократно повторяющийся мотив AUUUA, который и является элементом нестабильности. Сходная последовательность была обнаружена в 3'-НТО мРНК для другого онкобелка - c-fos. Однако в этой мРНК имеется еще один элемент нестабильности, который находится в транслируемой области.
Содержание свободного железа в клетке зависит не только от количества в клетке белка ферритина, но и от скорости его поступления в клетку. Скорость поступления железа в клетку возрастает с увеличением на поверхности клетки количества рецепторов для белка трансферина. Последний выполняет функцию переносчика железа. Оказалось, что синтез рецептора трансферина, так же как и ферритина, регулируется железом, но совсем по другому механизму. В присутствии железа в клетке мРНК рецептора трансферина быстро деградирует и синтез этого белка замедляется, в то время как при дефиците железа внутри клетки эта мРНК стабилизируется, обеспечивая высокий уровень синтеза белка. Нестабильность мРНК рецептора трансферина в присутствии железа определяется пятью шпилечными структурами в 3'-НТО (рис. 4, б ). Эти шпилечные структуры (элементы нестабильности мРНК рецептора трансферина) очень похожи на регуляторный элемент в 5'-НТО мРНК ферритина. Элементы нестабильности мРНК рецептора трансферина, так же как регуляторный элемент мРНК ферритина, связывают аконитазу в форме, не содержащей железа. В комплексе с аконитазой мРНК стабилизируется и активно транслируется. В результате на поверхности клеток увеличивается количество рецепторов трансферина и повышается поступление железа в клетки. При увеличении содержания свободного железа в клетках оно связывается с аконитазой. Аконитаза диссоциирует из комплекса с мРНК. Молекула мРНК дестабилизируется и быстро распадается. Это ведет к снижению скорости синтеза рецептора трансферина, уменьшению количества рецепторов на поверхности клетки и замедлению поступления железа в клетку.
Таким образом, один и тот же клеточный белок - аконитаза выполняет в клетке различные функции: в железосодержащей форме катализирует одну из реакций цикла Кребса - обратимое превращение лимонной кислоты в изолимонную, в форме без железа связывается с регуляторным элементом в 5'-НТО мРНК ферритина и репрессирует ее трансляцию, а также с элементами нестабильности в 3'-НТО мРНК рецептора трансферина и защищает эту мРНК от деградации.
СПЕЦИФИЧЕСКИЕ ПОСЛЕДОВАТЕЛЬНОСТИ В мРНК ОТВЕЧАЮТ ЗА ЕЕ ВНУТРИКЛЕТОЧНУЮ ЛОКАЛИЗАЦИЮ
Разные мРНК в эукариотических клетках распределяются в них неодинаково, что особенно легко наблюдать в крупных зародышевых клетках. Наиболее хорошо изучено распределение мРНК в ооцитах плодовой мушки дрозофилы . Так, мРНК, синтезируемые на генах oskar, nanos и bicoid, несколько раз меняют положение в клетке после своего синтеза, что контролируется комплексом генов, пока, наконец, не займут своего окончательного положения: мРНК oskar и nanos на заднем, а мРНК bicoid на переднем конце яйца. Правильное положение этих мРНК задается специфическими последовательностями в их 3'-НТО. Интересно, что сигналы внутриклеточной локализации в 3'-НТО очень протяженные. Вероятно, они формируют сложные вторичные структуры. За окончательную локализацию мРНК oskar и bicoid отвечает белок - продукт гена staufen, узнающий специфическую двухспиральную структуру в 3'-НТО этих мРНК. Интересно, что активная трансляция мРНК oskar и bicoid происходит только в местах их специфической локализации, а их делокализация приводит к подавлению трансляционной активности за счет связывания со специфическими репрессорными белками.

Рис. 5. Схема расположения функциональных участков на молекуле мРНК
Таким образом, на примере развития дрозофилы видно, что регуляция активности различных мРНКможет задаваться их локализацией в разных клеточных компартментах. Продукты трансляции этих мРНК, вероятно, создают в клетке белковые градиенты, и это, в свою очередь, лежит в основе дифференцировки клеток и определяет очередность трансляции и активность различных мРНК в процессе развития. Процесс, определяющий локализацию мРНК в клетке, как правило, состоит из нескольких последовательных стадий, обеспечивающих транспорт новосинтезированных молекул из ядра и затем "заякоривание" их в цитоплазме. Так, транспорт мРНК гена ug1 дрозофилы после ее выхода из ядра осуществляется путем взаимодействия с тубулином микротрубочек , тогда как "заякоривание" этой мРНК происходит в результате ее связывания с актиновыми филаментами .
ЗАКЛЮЧЕНИЕ
Таким образом, в молекулах мРНК содержится информация не только о последовательности аминокислот для определенного белка или белков, но также информация о том, когда, в каком количестве, в каком месте клетки и при каких условиях этот белок будет синтезирован. Схема расположения соответствующей информации на молекуле мРНК показана на рис. 5. Последовательность аминокислот в белке закодирована в виде линейной последовательности нуклеотидов. Другая информация в мРНК может содержаться как в виде определенных нуклеотидных последовательностей, так и в форме определенных пространственных структур, которые образует мРНК. Информация об одном и том же свойстве мРНК может содержаться в разных частях молекулы мРНК, иногда отстоящих друг от друга вдоль полинуклеотидной цепочки на значительном расстоянии. Весьма вероятно, однако, что эти участки молекулы мРНК сближаются при формировании ее пространственной структуры. Функциональные сигналы в мРНК могут узнаваться рибосомами, молекулами РНК (тРНК, регуляторными РНК) или белками.
Благодарю моих коллег В.М. Евдокимову, Д.В. Нащекина и П.В. Рузанова за ценные советы и помощь при подготовке статьи.
Список литературы
1. Спирин А.С. Молекулярная биология: Структура рибосомы и биосинтез белка. М.: Высш. шк., 1986.
2. Спирин А.С. Регуляция трансляции мРНК-связывающими факторами у высших эукариот//Успехи биол. химии. 1996. Т. 36. С. 3-48.
3. Высоцкая В.С., Гарбер М.Б. Регуляция экспрессии генов рибосомных белков Escherichia coli // Там же. 1995. Т. 35. С. 67-95.