Статья: Молекулярные машины
Обычно курс биологии строится от изучения простого, составных частей, к все более сложному. Сначала изучают химический состав клеток; потом ДНК, РНК, белок; затем строение клетки.
Но начать мы решили с чего-то более близкого к человеку с физическим образованием. Честно говоря, когда я изучала биологию, эта тема меня просто поразила, она мне показалось одной из наиболее интересных. Поэтому я решила вас не томить, не откладывать такую интересную тему на потом, а начать наш курс с рассмотрения работы молекулярных машин. Сегодня мы рассмотрим некоторые молекулярные машины. Первая из них называется АТФ-синтаза. Она занимается в митохондриях синтезом аденозинтрифосфорной кислоты (АТФ). Напомню, что АТФ – это молекула, которая обеспечивает клетку энергией (рис. 5).
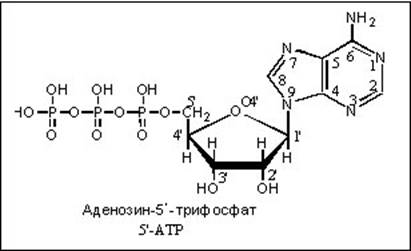
Для нас сейчас важно, зато молекула аденозинтрифосфорной кислоты содержит так называемую макроэргическую связь. Реакция синтеза представлена на схеме.
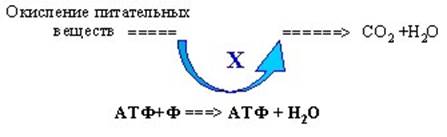
АДФ+Ф ==> АТФ +H2O
Из аденозиндифосфата и фосфата получается АТФ, при этом образуется так называемая макроэргическая связь, и на ее образование затрачивается 30,6 кДж/моль (7,3 ккал/моль). АТФ обеспечивает энергией большинство происходящих в клетке процессов, так как при гидролизе макроэргической связи запасенная в ней энергия освобождается.
Как же синтезируется эта молекула, то есть, как образуется макроэргическая связь между фосфатами? Это было одно время загадкой. Существовало предположение о том, что есть какое-то вещество Х, химический посредник, осуществляет связь между процессами, дающими энергию, то есть окислением питательных веществ до СО2 и Н2О, и каким –то образом энергия окисления (в своем роде медленное "горение" внутри организма) переходит в энергию макроэргической связи в молекуле АТФ. Это предположение о наличии химического посредника, которого никто найти не мог, называлось гипотезой химического сопряжения (рис. 6).
Но в 1961 г. английский ученый Питер Митчелл предложил другое объяснение – хемиосмотическую гипотезу (подробнее мы о ней будем говорить позже), которая заключается в том, что вода, которая образуется в процессе окисления, образуется не в виде молекулы воды, а виде протона H+ и иона гидроксила OH–. Энергия, получаемая при окислении, идет на то, чтобы продукты реакции – протон и гидроксил – разделить в пространстве. Протон выбрасывается из митохондрий через внутреннюю мембрану в межмембранное пространство (сам по себе протон не может проникнуть через мембрану митохондрии, эта мембрана непроницаема для заряженных частиц), и гидроксогруппы, которая остается внутри митохондрии.
В результате возникает разница концентраций ионов водорода (∆рН – то есть кислотности среды) и разница потенциала: положительные заряды снаружи митохондриальной мембраны, а отрицательный внутри. Напомним, что у митохондрий 2 мембраны, причем внешняя в энергетических процессах такой важной роли, как внутренняя, не играет. То есть энергия, полученная при окислении, запасена в виде электрохимической энергии. Электрический потенциал на мембране митохондрий достигает 200 милливольт, а толщина мембраны не превышает 10 нм.
Питер Митчелл первый высказал предположение о том, что химические реакции в клетке пространственно упорядочены, и продукты реакции распределяются асимметрично: протон в одну сторону, гидроксил в другую. За счет этого появляется электрохимический потенциал на мембране (обозначается Δμн). Он состоит из химической (∆рН – разница в концентрации протонов) и электрической (Δφ – разница в величине заряда) компоненты Δμн=∆рН + Δφ. Электрохимический потенциал на мембране митохондрий – универсальная форма запасания энергии клеткой.
Протоны могут перекачиваться через мембрану и при фотосинтезе в хлоропластах или в клетках фотосинтезирующих бактерий (Рис. 8).
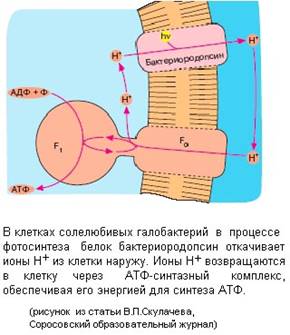
На рисунке представлена довольно простая система бактериального фотосинтеза, сопряженного с синтезом АТФ на примере галобактерий. Галобактерии живут в Мертвом море. Море настолько соленое, что соль выпадает в осадок, но в таких экстремальных условиях галобактерии прекрасно себя чувствуют. Галобактерии используют фотосинтез для получения энергии. Белок бактериородопсин под действием света выкачивает протоны изнутри бактериальной клетки наружу, и на мембране снаружи избыток протонов, и, соответственно, образуется положительный заряд. То есть в данном случае электрохимический потенциал на мембране бактерии возникает не за счет окисления веществ в процессе дыхания, а за счет работы, связанной со световой энергией.
Если протон "падает" сквозь мембрану внутрь митохондрии, при этом его потенциальная энергия уменьшается, так как он "падает" в электрическом поле от положительного заряда к отрицательному, и вдобавок по градиенту концентрации. Эта энергия используется для синтеза АТФ. И далее пойдет речь о том, как это происходит.
Синтезом АТФ занимается молекулярная машина, которая называется АТФ-синтаза. Она состоит из двух частей. Первая погружена в мембрану называется F0 (см. рисунок). Она представляет собой протонный канал, то есть это дыра в мембране, по которой протон может попасть внутрь митохондрии, но попадает он внутрь с потерей энергии, которую улавливает вторая часть молекулярной машины, которая называется F1. Эта часть АТФ-синтазы торчит внутрь митохондрии и использует энергию "падающих" через F0 протонов для того, чтобы аденозиндифосфат соединился с фосфатом посредством макроэргической связи и образовал молекулу АТФ.
Рассмотрим, как АТФ-синтаза синтезирует АТФ. Оказывается, что прежде всего совершается работа механическая, так как для осуществления синтеза АТФ в АТФ-синтазе крутится белковая структура. Как устроена АТФ-синтаза?
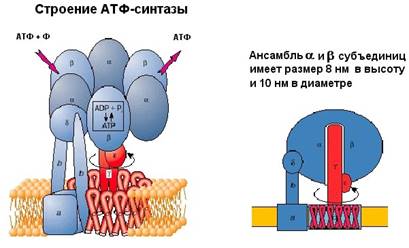
Она состоит из двух частей – статора (на рисунке 9 помечено синим цветом), и ротора (обозначен красным). Статор состоит из трех альфа субъединиц и трех бета субъединиц – они занимаются химической частью работы: синтезом АТФ из АДФ и фосфата. В собранном состоянии все вместе эти субъединицы по форме напоминают слега приплюснутый шар 8 нм в высоту и 10 нм в диаметре.
К ним примыкает дельта субъединица, и все вместе эта система образует F1 субъединицу молекулярной машины. Здесь же есть опора, которая «якорит» всю систему в мембране. Как известно, мембрана сделана из фосфолипидов (на рисунке показаны желтым). Гидрофильные "головки" фосфолипидов обращены в водную поверхность, а гидрофобные "хвосты" погружены внутрь мембраны, и именно они препятствуют перемещению заряженных частиц через мембрану. Вращающаяся часть машины, ротор, состоит из гамма и эпсилон субъединиц. Эта конструкция погружена в структуру, сделанную из одинаковых белков, они обозначаются буквой с. Статор держится в мембране, а ротор крутится. И энергия протона используется на то, чтобы прокрутить ротор этой машины.
Молекулярная машина работает в обе стороны (так же как и катализаторы, которые проводят реакцию как в прямую, так и в обратную стороны). Если течет протонный ток с наружной мембраны внутрь, то синтезируется АТФ; если же протонного потенциала нет, но подать с внутренней стороны АТФ, то машина начнет «выкачивать» протоны, создавая протонный потенциал. При этом ротор также вращается.
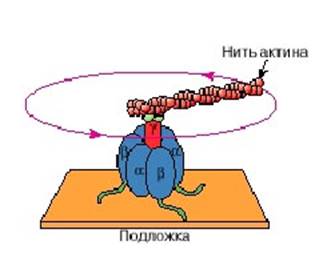
Для того, чтобы доказать, что в АТФ синтазе вращается часть машины, F1 фрагмент перевернули, «пришили» к неподвижной подложке, а к гамма-субъединице навесили искусственным образом нить актина (длинный белок, который можно было увидеть в микроскоп, так как он был мечен флуоресцентной меткой). Затем подали к этой системе энергию в виде АТФ, и оказалось, что при наличии АТФ гамма субъединица начала крутиться. Все это сняли на пленку. Было видно, как крутится флуоресцентная метка на актиновом хвостике, и было показано, что действительно происходит вращение во время работы этой молекулярной машины (рис. 10).
Теперь разберем, как же крутится этот ротор; как работает электромотор в мембране клеток, как у бактерий, так и у митохондрий высших организмов. Если вы вспомните временную ось возникновения жизни, то увидите, что возникнуть этот мотор должен был более трех миллиардов лет назад.
Как же используется протонный ток, чтобы крутить мотор? Оказалось, что в статоре имеется протонный канал, т.е. такой белок, который образует проход для протона. Но этот канал не сплошной. Если бы был канал, который пронизывал всю мембрану насквозь, то из-за разницы потенциалов все протоны потекли бы внутрь митохондрии, и произошла бы деэнергетизация мембраны, т.е. она бы разрядилась. Но канал устроен очень хитро. Он состоит из двух половинок (полу-каналов), которые, к тому же, смещены одна относительно другой (рис. 1).
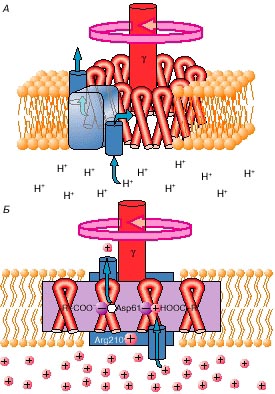
Структура этой машины такова, что протон проваливается через полуканал с наружной стороны митохондриальной мембраны, но попасть внутрь митохондрии он не может. Сваливается протон на подставленную ему аминокислоту ротора и эту аминокислоту протонирует, то есть на аминокислоте появляется дополнительный положительный заряд. Затем, когда протонированная аминокислота на вращающемся роторе доедет до следующей половинки канала, ведущей уже внутрь митохондрии (а внутри протонов мало и, кроме того, там протон поджидают отрицательно заряженные ионы), то протон наконец "падает" внутрь и аминокислота освобождается от положительного заряда. Заряды в роторе и статоре расположены таким образом, что протонирование – депротонирование приводит к повороту машины. Таким образом, протон в два приема проваливается внутрь митохондрии, и за счет этого мотор проворачивается.
За объяснение ферментативного механизма, лежащего в основе синтеза АТФ, два исследователя получили Нобелевскую премию: Пол Д. Бойер, США и Джон Э. Уолкер, Великобритания (Нобелевская премия 1997 года).
Было рассказано, как мотор крутится, но не было объяснено, почему синтезируется АТФ. Сейчас подробно мы на этом останавливаться не будем, но вкратце, объяснить это можно следующим образом. Представим АТФ в таком виде: АТФ=АДФ~Ф. Собственно, почему при разрыве этой связи выделяется большое количество энергии? При разрыве образуется отрицательно заряженный фосфат, который гидратируется (покрывается «шубой» из молекул воды). Как вы помните, вода – это диполь (кислород имеет частично отрицательный заряд, а два водорода - положительный). И за счет гидратирования эта энергия и получается. Но если синтез АТФ идет в той среде, где воды нет, т.е. в гидрофобной среде, то макроэргической эта реакция не является. Показано, что когда происходит образование ковалентной связи между фосфатными группами молекул АДФ и Ф, ферменту практически не требуется энергии. Реакции синтеза и гидролиза ATP в каталитическом центре фермента активно идут при отсутствии внешнего источника энергии. Условия, в которых находятся молекулы АДФ и Ф в каталитическом центре, существенно отличаются от условий протекания реакции в водной среде, благодаря чему образование молекулы АТФ в активном центре фермента может происходить энергетически "бесплатно". Энергия "падающих" протонов тратится потом на то, чтобы «выпихнуть» вон АТФ, отцепить его от каталитической субъединицы.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--