Статья: Получение мембранного белка бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот L-фенилаланина, L-тирозина и L-триптофана
Московская государственная академия тонкой химической технологии им. М.В. Ломоносова, 117571, Москва, проспект Вернадского, д. 86.
Биосинтетически получены препараты мембранного белка бактериородопсина, в которых природные ароматические аминокислоты L-фенилаланин, L-тирозин и L-триптофан селективно замещены на их дейтерий-меченные аналоги. Это достигается путем культивирования штамма галофильных бактерий Halobacterium halobium ЕТ 1001 на синтетической среде, содержащей вместо природных аминокислот химически синтезированные L-[2,3,4,5,6-2 H5 ]-фенилаланин, L-[3,5-2 H2 ]-тирозин и L-[2,4,5,6,7-2 H5 ]-триптофан. Представлены данные по культивированию H. halobium ЕТ 1001 на средах, содержащих дейтерий-меченные аналоги ароматических аминокислот и выделению дейтерий-меченного бактериородопсина. Анализ степени дейтерированности аминокислот гидролизатов бактериородопсина проводили методом масс-спектрометрии электронного удара после препаративного разделения их методом обращенно-фазовой высокоэффективной жидкостной хроматографии (ВЭЖХ) в виде метиловых эфиров дансил-аминокислот и бензилоксикарбонильных производных аминокислот. Показано, что в масс-спектрах производных аминокислот гидролизатов бактериородопсина присутствуют молекулярные ионы, которые соответствуют ароматическим дейтерий-меченным аминокислотам и практически отсутствуют их нативные немеченные аналоги. Полученные данные свидетельствуют о высокой эффективности мечения бактериородопсина в этих условиях.
ВВЕДЕНИЕ.
Ретинальсодержащий белок - бактериородопсин, выполняющий функции АТФ-зависимой транслоказы в клеточной мембране галофильных бактерий Halobacterium halobium , был выделен и детально проанализирован Остерхельтом в 1970 году [1]. Несмотря на то, что бактериородопсин в настоящее время довольно хорошо изучен, он все еще остается в центре внимания исследователей по целому ряду причин. Прежде всего, благодаря своей высокой светочувствительности и разрешающей способности, он широко используется в прикладных целях как биологический фотохромный материал [2]. Кроме этого, бактериородопсин весьма привлекателен, как модельный объект для исследований, связанных с изучением функциональной активности и структурных свойств мембранных белков в составе нативных энергопреобразующих мембран.
Компьютерная модель мембранного белка бактериородопсина из H. halobium
Для получения детальной информации о структуре полипептидной цепи мембранного белка в нативной мембране целесообразно селективно вводить в белок изотопные метки, которые позволяют использовать спектральные методы высокого разрешения, такие как спектроскопия ядерного магнитного резонанса (ЯМР) [3], Раман и лазерная спектроскопия [4,5], инфракрасная (ИК) спектрометрия [6] и масс-спектрометрия (МС) [7]. В связи с этим особенно перспективны исследования с мембранными белками, селективно обогащенными стабильными изотопами, например, дейтерием по остаткам таких функционально важных аминокислот, как L-фенилаланин, L-тирозин и L-триптофан [8,9]. Это связано с тем, что указанные аминокислоты участвуют в процессе формировании хромофорного центра бактериородопсина и в его функционировании [10]. Поэтому важно получать подобные модифицированные дейтерием белки в очищенном виде и в препаративных количествах.
Целью настоящей работы было получение препаратов бактериородопсина, селективно меченных дейтерием по остаткам ароматических аминокислот - L-фенилаланина, L-тирозина и L-триптофана, а также масс-спектрометрический анализ дейтерий-меченных аминокислот в составе гидролизатов бактериородопсина после их препаративного разделения методами обращенно- фазовой высокоэффективной хроматографии (ВЭЖХ) в виде метиловых эфиров дансил-аминокислот и бензилоксикарбонильных производных аминокислот.
УСЛОВИЯ ЭКСПЕРИМЕНТА .
В работе использовали соли квалификации “х.ч”., DL-аминокислоты (Reanal, Венгрия), аденозин-и уридин-5-монофосфаты (Sigma, США.), панкреотическую телячью дезоксирибонуклеазу 1 (Fluka Chemie AG, Швейцария), додецилсульфат натрия (ДСН) (Chemapol, Чехо-Словакия). L-[2,3,4,5,6-2 H5 ]-фенилаланин (90 ат.% 2 Н), L-[3,5-2 H2 ]-тирозин (96 ат.% 2 Н) и L-[2,4,5,6,7-2 H5 ]-триптофан (98 ат.% 2 Н) (способы получения указаны в работах [ 11,12]), были предоставлены доц. кафедры биотехнологии Московской государственной академии тонкой химической технологии им. М.В. Ломоносова к.х.н. А.Б. Пшеничниковой. Для получения производных аминокислот использовали N-диметиламинонафталин-5-сульфохлорид (дансилхлорид) (Sigma, США), карбобензоксихлорид (Войковский химзавод, РФ) и диазометан. Диазометан получали из N-нитрозометилмочевины (Merck, Германия).
Бактериальные штаммы и питательные среды . Объектом исследования служил пигментированный штамм галофильных бактерий Halobacterium halobium ЕТ 1001. Штамм был получен из коллекции культур микроорганизмов Московского государственного университета им. М.В. Ломоносова. Штамм поддерживали на пептоновой среде с 2%-ным агаром [13].
Для получения немеченного бактериородопсина использовали синтетическую среду, содержащую 18 аминокислот (количества компонентов приведены в г/л): (DL-аланин 0,43, L-аргинин 0,4, DL-аспарагиновая кислота 0,45, L-цистеин 0,05, L-глутаминовая кислота 1,3, L-глицин 0,06, DL-гистидин 0,3, DL-изолейцин 0,44, L-лейцин 0,8, L-лизин 0,85, DL-метионин 0,37, DL-фенилаланин 0,26, L-пролин 0,05, DL-серин 0,61, DL-треонин 0,5, L-тирозин 0,2, DL-триптофан 0,5, DL-валин 1), нуклеотиды (аденозин-5-монофосфат 0,1, уридин-5 монофосфат 0,1), соли (NaCl 250, MgSO4 7H2 O 20, KСl 2, NH4 Cl 0,5, KNO3 0,1, KH2 PO4 0,05, K2 HPO4 0,05, цитрат натрия 0,5, MnSO4 H2 O 3 10-4 , CaCL2 6H2 O 0,065, ZnSO4 7H2 O 4 10-5 , FeSO4 7H2 O 5 10-4 , CuSO4 5H2 O 5 10-5 ), глицерин 1, ростовые факторы (биотин 0,1 10-3 , фолиевая кислота 10 10-3 , витамин В12 0,02 10-3 ).
В экспериментах по введению дейтериевой метки в бактериородопсин вместо L-фенилаланина, L-тирозина и L-триптофана в синтетическую среду добавляли их дейтерированные аналоги - L-[2,3,4,5,6-2 H5 ]-фенилаланин, L-[3,5-2 H2 ]-тирозин, и L-[2,4,5,6,7-2 H5 ]-триптофан.
Среды стерилизовали при 0,5 ати в течении 30-40 мин, рН доводили до величины 6,5-6,7 при помощи KOH. Посевной материал выращивали в колбах Эрленмейера, объемом 250 мл (с наполнением средой до 50 мл) при 35-37 0 С в условиях интенсивной аэрации и освещении лампами дневного света ЛДС-40. После суток посевной материал в количестве 5-10 % переносили в синтетическую среду, содержащую дейтерий-меченные аналоги ароматических аминокислот и культивировали в течении 4-5 суток как и при получении посевного материала.
Выделение фракции пурпурных мембран . Клетки осаждали на центрифуге Т-24 (Германия) (10000 об/мин, 10 мин.), осадок клеток суспендировали в дист. воде, экспонировали ультразвуком (3 экспозиции продолжительностью 5 мин) и вновь центрифугировали (10 000 об/мин, 10 мин). Пигменты удаляли обработкой ацетоном, липиды экстрагировали смесью хлороформ-метанол (2:1), растворитель декантировали. Полученный осадок клеток (100-150 мг) суспендировали в 100 мл буфера трис-HCL (рН 7,5), добавляли 1 мг дезоксирибонуклеазы 1 и инкубировали в течении 5-6 часов при 37 0 С, затем разбавляли дист. водой до 200 мл, после чего инкубировали 15 часов при 40 С. Осадок промывали водой с последующим отделением растворителя до получения бесцветных промывных вод. Контроль чистоты полученной суспензии пурпурных мембран (в Н2 О) проводили на спектрофотометре “Beckman DU-6” (США) по соотношению полос поглощения при 280 нм/568 нм (e280 =1,1 10 5 М-1 см-1 [14] и e568 =6,3 104 М-1 см-1 [15]).
Выделение бактериородопсина . Препараты пурпурных мембран (50 мг) солюбилизировали в 2 мл 0,5%-ного раствора додецилсульфата натрия (ДСН) в Н2 О, выдерживали в течении 8-10 часов при 220 С, затем центрифугировали (7000 об/мин, 5 мин). Осадок отделяли, к супернатанту добавляли 5-ти кратный избыток метанола, выдерживали при 00 С в течении 12-14 часов и центрифугировали (13000 об/мин, 10 мин). Остатки ДСН удаляли, промывая осадок белка дист. водой. Выход бактериородопсина составил 20 мг.
Гидролиз бактериородопсина . Гидролиз бактериородопсина проводили двумя методами: (А)-кислотным и (В)-щелочным гидролизом. Для этого белок делили на две равные порции по 10 мг и гидролизовали в запаянных стеклянных ампулах в 30-ти кратном избытке гидролизирующего агента (24 ч, 1100 С): (А)- в 6 н. 2 НCL (в 2 Н2 O ) с 3%-ным (по весу) фенолом и в (В) - 4 н. Ba(OH)2 . После этого гидролизат, полученный в условиях (А) упаривали в роторном испарителе при 400 С. Остатки дейтеро-соляной кислоты удаляли путем выдерживания в эксикаторе над твердым NaOH. После проведения гидролиза (В), реакционную смесь суспендировали в одном объёме горячей дистиллированной воды и нейтрализовали 2 н. раствором H2 SO4 до рН 7,0. Выпавший осадок сульфата бария отделяли центрифугированием (10000 об/мин, 5 мин), супернатант декантировали и упаривали в роторном испарителе при 400 С. Гидролизаты бактериородопсина, полученные в условиях (А) и (В), высушенные до постоянной массы, обрабатывали дансилхлоридом и диазометаном (или карбобензоксихлоридом).
Получение дансиламинокислот гидролизатов бактериородопсина . К 10 мг высушенных гидролизатов бактериородопсина в 2 мл 2 м. NaHCO3 (2 10-4 моль) рН 9-10 дробными порциями при перемешивании добавляли 16 мг (5,9 10-5 моль) дансилхлорида в 4 мл ацетона. Реакционную смесь выдерживали при перемешивании при 400 С в течении часа, затем подкисляли 2 н. раствором HCL до рН 3 и экстрагировали этилацетатом (3 раза по 5 мл.). Объединенный экстракт промывали дист. водой до значения рН 7,0, сушили безводным сульфатом натрия, растворитель удаляли при 10 мм. рт. ст.
Получение метиловых эфиров дансиламинокислот гидролизатов бактериородопсина . К 20 мл 40 %-ного КОН в 40 мл эфира добавляли 3 г влажной нитрозометилмочевины и перемешивали на водяной бане со льдом в течении 15-20 мин. После интенсивного газовыделения эфирный слой отделяли и промывали ледяной водой до рН 7,0, сушили безводным NaSO4 и обрабатывали им препараты дансилпроизводных аминокислот в составе гидролизатов бактериородопсина.
Бензилоксикарбонильные производные аминокислот получали реакцией Шоттен-Баумана по методике, указанной в работе [16].
Тонкослойную хроматографию (ТСХ) производных аминокислот проводили на пластинках “Silufol UV-254” (Чехо-Словакия) в системах: (A)-хлороформ-метанол-уксусная кислота (10:1:0,3) для бензилоксикарбонильных производных аминокислот и (Б)-хлороформ-метанол-ацетон (7:1:1) для метиловых эфиров дансиламинокислот. Аналитическое и препаративное разделение карбобензоксипроизводных аминокислот и метиловых эфиров дансиламинокислот в составе гидролизатов бактериородопсина проводили методом обращенно-фазовой высокоэффективной жидкостной хроматографии (ВЭЖХ) на жидкостном хроматографе “Knauer” (ФРГ), снабженным насосом “Knauer”, УФ-детектором “2563” и интегратором “С-R 3A” (Shimadzy, Япония). Использовали колонки: гиперсил ODS, 5 мкм, 3 250 мм, силасорб С18, 12 мкм, 10 250 мм. Элюирование проводили в системе растворителей по двум вариантам: (А)- вода-трифторуксусная кислота (100/0,1-0,5 об/об) и (В)- ацетонитрил-трифторуксусная кислота (100/0,1-0,5 об/об) при различных градиентных режимах, как описано в работе [17]. Бензилоксикарбонильные производные аминокислот детектировали по поглощению при 254 нм. Метиловые эфиры дансил-аминокислот детектировали по флуоресценции в УФ-свете.
Масс-спектры электронного удара производных аминокислот получены на приборе “MB-80 A” (Hitachi, Япония) при энергии ионизирующих электронов 70 эВ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ .
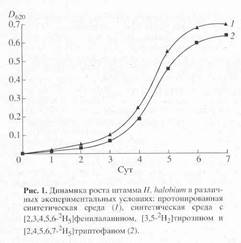
Получение дейтерий-меченного бактериородопсина . Выбор способа получения бактериородопсина был определен целью исследования, связанной с изучением принципиальной возможности получения дейтерий-меченных препаратов бактериородопсина в препаративных количествах. При выборе L-[2,3,4,5,6-2 H5 ]-фенилаланина, L-[3,5-2 H2 ]-тирозина и L-[2,4,5,6,7-2 H5 ] -триптофана в качестве источников изотопных меток, авторы учитывали важность этих аминокислот в функционировании бактериородопсина, стабильность дейтерий-меченных аналогов указанных аминокислот к (1 H-2 H) обмену в водной среде в условиях культивирования, а также возможность их детектирования методами масс-спектрометрии. Кривые, отражающие динамику роста штамма H. halobium ЕТ 1001 на пептоновой среде (1), обычной синтетической среде(2) и синтетической среде (3), содержащей дейтерий-меченные аналоги аминокислот L-Phe, L-Tyr и L-Trp представлены на рис. 1. Как видно из рис.1, рост H. halobium ЕТ 1001 на пептоновой среде происходит лучше чем на синтетической, однако для получения дейтерий-меченного бактериородопсина пептоновая среда не подходит ввиду наличия в ней немеченных аминокислот. Поэтому пептоновая среда была заменена на синтетическую среду, в которую добавляли дейтерий-меченные аналоги ароматических аминокислот.
Способ получения дейтерий-меченного бактериородопсина представлен на схеме. Основными этапами при получении дейтерированных препаратов бактериородопсина было: культивирование штамма H.halobium ЕТ 1001 на средах, содержащих дейтерий-меченные аналоги ароматических аминокислот, разрушение клеток и получение суспензии пурпурных мембран, очистка суспензии пурпурных мембран от обломков клеток и клеточной ДНК, очистка от каротиноидов, солюбилизация белка в додецилсульфате натрия (ДСН) и осаждение конечного продукта метанолом (см. схему). При получении пурпурных мембран, клетки обрабатывали дезоксирибонуклеазой, для того чтобы разрушить клеточную ДНК. Вследствие того, что препараты пурпурных мембран содержат примеси каротиноидов и обломки клеточных стенок, было необходимо применять специальные методы очистки и выделения белка. При выделении фракции белков из бактериальных объектов необходимо учитывать наличие в них небелковых примесей. В случае богатых белками штаммов сравнительно небольшим количеством примесных соединений в них часто пренебрегают, используя в качестве белковой фракции остаток после исчерпывающего отделения пигментов и липидов экстракцией органическими растворителями. В специальных случаях для получения фракции индивидуальных белков следует прибегать к их осаждению и очистке [18]. Выделение мембранного белка заключается в его солюбилизации в растворе детергента и последующим осаждении белка. Для этого пурпурные мембраны суспендировали в растворе ДСН. Выбор ДСН в качестве детергента в данном случае оправдан, так как бактериородопсин является интегральным белком. Выделение бактериородопсина после отделения осколков клеточных стенок из раствора ДСН достигали, осаждая белок метанолом. Наиболее трудоемкой была стадия очистки бактериородопсина от каротиноидов, которая приводила к значительным потерям хромопротеида. Для этого был использован метод низкотемпературной экстракции каротиноидов ацетоном.
![]() Культивирование H. halobium ЕТ 1001на синтетической среде, содержащей L-[2,3,4,5,6-2 H5 ] -фенилаланин, L-[3,5-2 H2 ]-тирозин и L-[2,4,5,6,7-2 H5 ]-триптофан.
Культивирование H. halobium ЕТ 1001на синтетической среде, содержащей L-[2,3,4,5,6-2 H5 ] -фенилаланин, L-[3,5-2 H2 ]-тирозин и L-[2,4,5,6,7-2 H5 ]-триптофан.
![]()
![]()
![]() Культуральная жидкость
Культуральная жидкость
![]() Отделение биомассы
Отделение биомассы
![]() Сырая биомасса
Сырая биомасса
![]()
![]() Промывка клеток дист. Н2 О
Промывка клеток дист. Н2 О
--> ЧИТАТЬ ПОЛНОСТЬЮ <--