Статья: Популяции. Дрейф генов
Группы особей одного вида, населяющие территории, разделенные естественными преградами (реками, горами, пустынями), являются относительно самостоятельными, поскольку скрещивание между ними затруднено. Такие группы называют популяциями (термин ввел в 1903 г. датский ученый В. Иогансен, для того чтобы отличать генетически разнородную группу особей от однородной чистой линии). Вид, таким образом, состоит из популяций, каждая из которых занимает часть ареала вида. За многие поколения в генофондах популяций накапливаются те аллели исходных популяций, которые обеспечивают наибольшую приспособленность к местам обитания. Вследствие различий в исходном генном составе и обитания в разных природных условиях генофонды изолированных популяций отличаются частотой встречаемости аллелей, поэтому один и тот же признак в разных популяциях может проявляться по-разному. Так, северные популяции млекопитающих имеют более густой мех, а южные — более темный.
Согласно Писанию, организмы, как и сама планета, созданы Творцом. Однако их природа изменчива. Чтобы понять, какие основные группы сотворены, а какие могли появиться впоследствии, необходимо исследовать степень изменчивости организмов. Подтверждают ли наблюдаемые изменения гипотезу эволюции (лат. evolutio развертывание) — исторического саморазвития материи? Ученые различают микро- и макроэволюцию. Под микроэволюцией понимаются изменения в популяциях в ряду последовательных поколений вплоть до появления новых видов, происходящие за относительно короткий промежуток времени (иногда их наблюдают в природе). Под макроэволюцией подразумевают гипотезу длительного исторического саморазвития, приводящего к появлению надвидовых таксонов (родов, семейств, отрядов).
Рассмотрим два основных типа изоляции и обусловленные ими изменения.
Географическая изоляция
Организмы пространственно изолированных популяций могут развиваться различными путями. Особи разных популяций могут потерять возможность скрещиваться по причине возникших различий в сроках брачных периодов, в инстинктах сооружения гнезд и нор, в поведении в период спаривания. У особей может измениться брачная окраска (рыбы, птицы), брачные песни (птицы, земноводные, насекомые) или брачные танцы (птицы, рыбы, членистоногие). Растения разных популяций могут иметь различные периоды цветения, разные взаимоотношения с опыляющими насекомыми. Процесс расхождения признаков в популяциях носит название дивергенции. Причиной дивергенции при географической изоляции является как приспособление существ к разным условиям среды, так и различие в исходных генофондах (в новые популяции разные аллели генов попадают в разных соотношениях).
Популяции лиственницы сибирской заселили огромную территорию от Урала до Байкала. В зонах с наиболее суровыми климатическими условиями обитает вид лиственницы даурской. На Дальнем Севере произрастает вид мака с быстрым развитием коротких цветоносов и ранним цветением, не характерными для умеренных широт. Ученые предполагают, что эти виды развились от одного исходного, хотя, конечно же, этого никто не наблюдал, а проверить реальность подобных микроэволюционных изменений в большинстве случаев не представляется возможным.
Растение прострел широко распространено в Европе. У западной формы прострела листья тонкие, цветки поникшие. Восточная форма, обитающая в засушливых условиях, имеет более широкие листья и стоячие цветки, по которым вода стекает к корню. Между западным и восточным прострелом существует непрерывный ряд промежуточных форм, имеющих выраженный приспособительный характер.
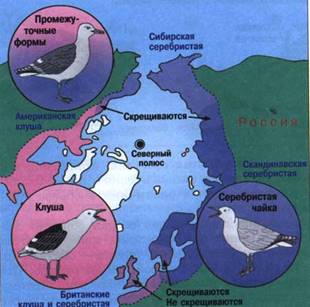
Вид синица большая представлен тремя подвидами, обитающими в трех группах ареалов: евроазиатской, южноазиатской и восточноазиатской. Южноазиатские синицы в южной и восточной зонах контакта скрещиваются с двумя другими подвидами. Восточноазиатские и евроазиатские, обитая совместно в долине Амура, не скрещиваются. Ученые предполагают, что подвиды синиц сформировались в четвертичный период с наступлением оледенения.
С оледенением связано, по мнению ученых, и возникновение родственных видов ландыша. Единый ареал вида, возможно, был разорван ледником на несколько изолированных частей. Ландыш, переживший оледенение, вновь широко распространился по всей лесной зоне, образовав в Европе новый более крупный вид с широким венчиком, а на Дальнем Востоке — вид с красными черешками и интенсивным восковым налетом на листьях.
В средней полосе произрастает около 20 видов лютика. Они занимают различные места обитания — луга, леса, берега рек и озер и т.д. Ученые предполагают, что вследствие изоляции лютики обособились сначала в подвиды, а затем — в виды.
На Галапагосских островах обитают птицы-вьюрки. Каждый вид имеет свои особенности. Например, большой земляной вьюрок питается семенами растений и имеет толстый массивный клюв, а дятловый вьюрок длинным прямым клювом достает из-под коры насекомых.
Формирование новых видов, связанное с географической изоляцией (разобщенностью), эволюционисты называют аллопатрическим (греч. allos иной и patris родина) видообразованием.
Экологическая изоляция
Другая причина, приводящая к дивергенции видов, — различие экологических ниш. Различия, например, в питании ведут к различиям в образе жизни и разобщенности особей.
Вид окунь обыкновенный в крупных озерах образует две популяции. Одни окуни живут в прибрежной зоне, питаются мелкими животными и растут медленно, другие — обитают на большей глубине, питаются рыбой и икрой, растут быстро. Популяции форели нерестятся в устьях различных рек, поэтому спаривание между ними затруднено и развитие происходит изолированно (такую изоляцию можно также назвать и географической). Некоторые лососевые рыбы мечут икру раз в два года. В одно и то же нерестилище попеременно приходят на нерест разные популяции: одна — в четные годы, другая — в нечетные.
Виды летучих мышей различаются способом питания: одни — хищные, а другие плодоядные. Некоторые виды пираньи с мощными челюстями и острыми зубами питаются исключительно растениями. Панда похожа на бурого медведя, но питается в основном бамбуком, лишь изредка поедая животных. Среди видов крокодила одни охотятся на животных, другие питаются только рыбой, а некоторые ископаемые виды были растительноядными.
Пять видов синиц сформировались, как предполагают ученые, вследствие различной пищевой специализации. Синица хохлатая питается семенами хвойных деревьев, лазоревка добывает мелких насекомых из расщелин древесной коры, синица большая ловит крупных насекомых в городских садах и парках, а гаичка и московка — в лесах.
Близкие виды обыкновенных европейских бабочек-белянок — капустница, репница и брюквенница — питаются на личиночной стадии различной пищей. Гусеницы капустницы и репницы поедают только культурные виды крестоцветных, а брюквенницы — дикие. Вид черный дрозд в настоящее время существует в двух формах. Одни дрозды селятся вблизи жилья человека, а другие — в глухих лесах. Эти два подвида внешне неразличимы.
По мнению ученых, некоторые виды растений в поймах крупных рек образовали виды, дающие семена до или после разлива. Виды, считающиеся исходными, растут на незаливаемых местах и опыляются, как правило, во время разлива.
Образование новых видов в прежнем ареале эволюционисты называют симпатрическим (греч. sym вместе и patris родина) видообразованием. Предполагают, что симпатрическое видообразование может быть вызвано не только внешними условиями, но и изменением кариотипа. В результате в одном ареале возникают генетически изолированные популяции. В роде хризантем все виды имеют число хромосом, кратное 9: 18, 27... 90, в роде картофеля — кратное 12: 24, 48, 72. Предположение о симпатрическом видообразовании основано на морфологической близости возможных исходных видов и дочерних форм.
Справедливо заключить, что вариации признаков, а также относительно правдоподобные гипотезы видообразования ограничены пределами групп очень сходных существ.
Принято считать, что главное биологическое значение изменений в популяциях состоит в обеспечении жизнеспособности организмов при наиболее полном использовании пространства и пищевых ресурсов. По мнению православных ученых. способность к изменениям заложена Творцом и позволяет организмам расселяться по всей планете, заполнять различные экологические ниши в соответствии с Его планом, направленным не только лишь на обеспечение жизнеспособности организмов. Неповторимая красота каждого уголка нашей планеты, ее флоры и фауны, — тоже результат Его заботы о мире. По этим причинам ряд ученых указывают, что использование термина "эволюция" в значении саморазвития применительно к изменениям в популяциях не вполне правомерно.
Закон Харди—Вайнберга
Рассмотрим бесконечно большую изолированную популяцию, скрещивания в которой происходят случайно (панмиксия), новые мутации не возникают, а особи с разными генотипами имеют одинаковую приспособленность.
Если в такой идеальной популяции частота встречаемости аллеля А будет р, то частота аллеля а будет q= 1 - р. Предположим, что все особи одновременно сформировали гаметы. Встречаемости разных гамет также будут p и q. После скрещивания встречаемость доминантных гомозигот будет р2 = рр (обе гаметы должны нести аллель А), рецессивных гомозигот — q2, а гетерозигот — 2рq = рq + qр (от "папы" — А, от "мамы" — а и наоборот). Таким образом, справедлив закон Харди—Вайнберга: в идеальной популяции соблюдается неизменное соотношение между доминантными и рецессивными аллелями (соответственно между гомо- и гетерозиготами). В аналитическом виде:
(рА + qа) х (рА + qа) = (рА + qа)2 = р2АА + 2рqАа + q2аа =1
Генетическая структура популяции станет неизменной уже после первого панмиксного скрещивания, ведь после того как гены оказались в гаметах, абсолютно неважно, какова была доля гомозигот среди родителей. Если, к примеру, в исходной популяции все особи были гомозиготами, половина — доминантными, другая — рецессивными, то в F1: 1/4АА + 1/2Аа+ 1/4аа. Такая же структура F1 будет и в случае, если все родители гетерозиготны.
Закон был открыт в 1908 г. английским математиком Дж. Харди и независимо от него немецким врачом Г. Вайнбергом. Пользуясь законом Харди—Вайнберга, можно узнать, какой процент особей имеет рецессивный ген по какому-нибудь наследственному заболеванию. Пусть встречаемость заболевания 1 %, то есть q2=0,01, тогда q= 0,1, а р= 0,9. Встречаемость рецессивных носителей — 2рq= 0,18, то есть 18 % особей.
В природных популяциях действуют так называемые факторы эволюции: естественный отбор аллелей, мутации, изоляция, дрейф генов. Действие этих факторов нарушает равновесие, отвечающее закону Харди—Вайнберга.
Дрейф генов
Если сравнить генофонд популяции с кувшином, полным разноцветных горошин, то сокращение численности будет соответствовать случайной выборке горсти горошин (аналогичным образом далеко не все гаметы родителей реализуются в потомстве). В новой популяции будет иная цветовая пропорция горошин-генов, некоторых цветов вообще может не оказаться. Соотношение между встречаемостью гомо- и гетерозиготных особей изменится, произойдет дрейф генов — случайное ненаправленное изменение частот аллелей в ряду поколений. Наиболее отчетливо дрейф проявляется при резком сокращении численности популяции в результате пожара, засухи, морозов, а также при заселении частью особей новых территорий с образованием новой популяции.
В природе наблюдаются чередующиеся подъемы и спады численности особей — популяционные волны (тип дрейфа). Размножение хищников находится в тесной взаимосвязи с колебаниями количества белок, зайцев, мышей. У насекомых и однолетних растений наблюдаются сезонные популяционные волны.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--