Статья: Поведение и оседание актинул tubularia larynx (leptolida, tubulariidae)
Освобожденная актинула падает на дно и становится на аборальные щупальца, закрепляясь на субстрате концевыми нематоцистами (Hawes, 1955). Длина ее тела составляет 5—6 мм, длина организованных в венчик 10—12 аборальных щупалец в расслабленном состоянии 8 мм, а длина семи-девяти оральных щупалец, организованных в околоротовой венчик”—0,5 мм. Длина ротового конуса и длина чашечки с зачатком ножки одинаковы — 0,3 мм (рис. 2, а). Наряду с нормальными актинулами из гонофоров появляются и аномальные формы: мы наблюдали личинок с двумя ртами и соответственно двумя оральными венчиками щупалец, без ротового конуса и орального венчика щупалец, а также с частично редуцированным аборальным венчиком, состоящим из двух-трех щупалец (рис. 2, б—г).
Актинула может слабо передвигаться по дну, перебирая щупальцами как ногами, цепляясь ими за окружающие предметы. На этой стадии личинка проявляет фотокинетическую активность: ее движения активизируются при ярком освещении и замедляются в темноте. Направленный свет, однако, не воздействует на движение, т. е. фототаксис отсутствует. Актинулы не проявляют также ни гео-, ни схизотаксисов.
Таблица 1
Прикрепление актинул Tubularia larynx в секторах пяти типов с различной структурой субстрата на дне трех экспериментальных установок разных размеров (“Материал и методика”)
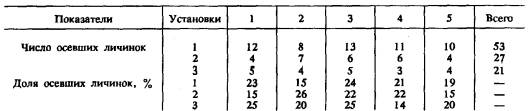
Примечание. Оседание актинул 100-процентное.
В экспериментальных сосудах с различной шероховатостью поверхности дна в разных секторах актинулы распределялись беспорядочно, не проявляя предпочтения ни к одному из секторов (табл. 1), что можно объяснить индифферентностью личинок к различным в структуре поверхности субстратам.
Течение воды, искусственно создаваемое в экспериментальной установке, практически не сказывалось на характере движения .и распределения актинул на ее дне. Актинулы способны противостоять водному потоку со скоростью до б см/с, если они заякорились только концевыми нематопистами щупалец за поверхность дна; личинки, зацепившиеся щупальцами за какие-либо предметы, способны выдерживать течение со скоростью до 12 см/с. Однако прикреплялись личинки лишь в стоячей воде или на течении со скоростью менее 5 мм/с. Следовательно, в море прикрепление, равно как и выход личинок из гонотек, происходят во время полной или малой воды. За период между освобождением и прикреплением мощные приливные или отливные течения разносят актинул по дну. Если же актинула сразу после выхода из гонофора успевает попасть в щель между камнями или в заросли обрастающих сообществ, где гидродинамика значительно слабее, она прикрепляется рядом с материнской колонией: так формируются уже описанные поля Т. larynx. Сила противостояния приливно-отливным течениям у актинул Г. larynx больше, чем у некоторых изученных гидроидов: так, планулы Gonothyraea Icweni способны к самостоятельному движению при скорости течения, не превышающей 4 мм/с; Laomedea flexuosa — 6 мм/с, Dynamena pumila — 9 мм/с; прикрепление планул указанных гидроидов происходит исключительно в неподвижной воде (собственные наблюдения).
Высокий предел скорости течения, при котором личинки могут оставаться в благоприятном местообитании, объясняет процветание Т. larynx на дне желоба в проливе Великая Салма, где встречаются лишь слаборазвитые колонии Abietinaria abietina (L.) (отряд Thecaphora).
Таблица 2
Оседание актинул Т. larynx в чашках Петри на слизистый бактериальный слой с поверхности разных субстратов

Примечание. Число первичных полипов выражено в процентах от общего количества личинок, выпущенных на пленку каждого типа. Представлены средние результаты трех серий экспериментов. Контроль — доля актинул, осевших на чистое стекло.
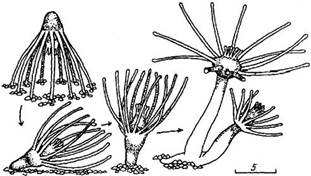
Рис. 3. Прикрепление актинулы и образование колонии. Масштаб 5 мм
Поверхностный бактериальный слой, стимулирующий оседание планул многих колониальных гидроидов (Scheltema, 1974; Chia, 1978), не активизирует прикрепление актинул T. larynx. В чашках Петри, дно которых было покрыто бактериальным слоем с разных погруженных субстратов, актинулы прикреплялись к поверхности дна, не проявляя предпочтения к какому-либо из использованных субстратов, в том числе и к стерильной поверхности (табл. 2).
Прикрепление
Через б—8 ч после выхода из гонофоров актинулы переворачиваются, держась нематоцистами щупалец за дно или окружающие тела (Hawes, 1955), и становятся на аборальный конец — подошву. Последняя выделяет слизистые вещества и перисарк в течение 3—4 ч” по истечении которых актинула превращается в прикрепленную форму — первичного полипа. Внешних морфологических изменений при этом не происходит, поэтому говорить о метаморфозе, свойственном другим колониальным гидроидным, в данном случае не приходится. При оседании на песок подошва актинулы склеивает несколько песчинок вокруг себя, формируя твердую опору для создания колонии (рис; 3).
Весь период жизнедеятельности свободнодвигающейся личинки от выхода из гонотеки до прикрепления занимает 12 ч. Заметим, что это время совпадает с приливно-отливным циклом в проливе Великая Салма. Это значит, что во время одной стоячей воды, например полной, актинулы освобождаются и оседают на дно; затем начинается отлив, во время которого часть актинул уносится в море, а другие, зацепившись или попав в различного рода укрытия, остаются рядом с материнской колонией. Через 12 ч — в малую воду личинки окончательно прикрепляются к субстрату, на котором они оказались в результате транспортировки течениями. Если место благоприятно для дальнейшего развития, первичный полип формирует колонию.
Ножка первичного полипа покрыта перисарком Стекой), появляющимся еще на стадии актинулы, с помощью которого аборальный конец личинки прикрепляется к субстрату (Hawes, 1958). Ножка полипа удлиняется, из его подошвы вырастает гидрориза, на которой начинается формирование второго гидранта. Начальная колония, состоящая из двух полноценных кормящихся гидрантов, образуется в естественных условиях через 60 ч после выхода из гонотек, что было выяснено при помощи подводного эксперимента в проливе на глубине 15 м.
Заключение
Таким образом, в результате наших исследований можно заключить, что:
— актинула Г. larynx выходит из гонотеки оральным концом во время спокойной воды;
— оседание актинул не определяется светом, свойствами субстрата или слизистым бактериальным слоем, покрывающим поверхность субстрата, т. е. место прикрепления определяется достаточно случайно;
— в первый же период спокойной воды, следующий за освобождением личинок, актинула прикрепляется к субстрату, на который ее занесло течением;
— процветание вица в местах с мощной гидродинамикой определяется способностью прикрепившихся актинул и формирующихся колоний выдерживать сильные приливно-отливные течения.
Список литературы
Наумов Д. В., 1980. Гидроиды, гидромедузы морских, солоноватоводных и пресных бассейнов СССР:Изд-во АН СССР. М. С. 1—585.
Орлов Д. В., Марфенин Н.Н., 1993. Поведение и оседание личинок Clava multicornis Forskal
(Ну drozoa)//Вести. МГУ. Сер. 16, №4. С. 24—30.
Chia F. S., 1978. Mechanisms of larval attachment and the induction of settlement and metamorphosis in Coelenlerales: a review//SetlIement and metamorphosis of marine invertebrate larvae (Chia F. S., Rice M. E.. eds.). New York. P. 1—12.