Дипломная работа: Распределение антигенов системы HLA у больных туберкулезом и здоровых представителей русской этнической группы Челябинской области
Актуальность данных исследований в первую очередь связана с необходимостью накопления информации в направлении "HLA и болезни" в русской этнической группе Челябинской области, а также для изучения ассоциации HLA с туберкулезом.
Новизна данного исследования заключается в том, что в процессе работы впервые был проведен анализ особенностей распределения специфичностей генов HLA II класса (DRB1, DQA1 и DQB1) и их гаплотипических сочетаний на молекулярно-генетическом уровне среди пациентов с туберкулезом и здоровых людей русской этнической группы, проживающих в Челябинской области. Иммуногенетический анализ больных туберкулезом в Челябинской области проводился впервые.
Объектом настоящего исследования являлись больные туберкулезом (51 человек), находящиеся на стационарном лечении в ГУЗ "Противотуберкулезный диспансер №3", представители социально благополучных слоев населения, имеющие родственников русской национальности в трех поколениях. Группу сравнения составили 202 случайно выбранных донора Областной станции переливания крови (г. Челябинск) русской национальности.
Цель работы: Выявить наличие ассоциации генов HLAII класса (локусов DRB1, DQA1 и DQB1) и их гаплотипов с развитием туберкулеза в русской этнической группе Челябинской области.
Задачи исследования:
1. Установить частоты встречаемости генов HLAII класса и их гаплотипов в группе больных туберкулезом.
2. Провести сравнение полученных данных с результатами распределения генов HLAII класса в контрольной группе.
3. Сделать вывод о наличии положительной или отрицательной ассоциации генов HLAII класса с развитием туберкулеза у представителей русской этнической группы Челябинской области.
Теоретической базой для настоящей работы являются популяционно-иммуногенетические исследования, проведенные ранее на других популяциях, а также литературные данные об ассоциации иммуногенетической системы HLA с развитием туберкулеза.
Генотипирование аллельных вариантов HLA II класса проводилось методом полимеразной цепной реакции. Для обработки результатов применялись стандартные генетико-статистические методы.
Практическая значимость работы:
1. Данные о распределении специфичностей генов HLA II класса и их сочетаний у здоровых представителей русской этнической группы Челябинской области могут быть использованы в качестве контрольных для поиска маркеров генетической предрасположенности к развитию различных заболеваний.
2. Полученные данные могут быть теоретически использованы для практических рекомендаций при лечении туберкулеза легких.
3. Результаты исследования ассоциации генов HLAIIкласса с развитием туберкулеза могут быть использованы в качестве дополнительного диагностического критерия при выявлении группы риска по данному заболеванию.
Композиция работы определяется ее целью и задачами и отражает основные этапы исследования.
I. Обзор литературы
1.1 Физиологическая роль главного комплекса гистосовместимости человека
Регуляция иммунного ответа является одной из основных физиологических функций организма. Эта функция принадлежит генам главного комплекса гистосовместимости человека. При этом следует принять во внимание, что само это название отражает скорее историю открытия данной генетической системы, чем ее основную функцию. Дело в том, что история открытия первых продуктов генов главного комплекса гистосовместимости человека, называемых антигенами HLA (от Human leucocyte antigens), связана именно с появлением и развитием трансплантационной иммунологии, когда возникла необходимость подбора тканесовместимых пар донор и реципиент. Сегодня же известно, что роль системы HLA в отторжении трансплантата является лишь одной из частных физиологических функций этой системы, а основная же ее функция - это регуляция иммунного ответа. В 80-х годах даже дискутировался вопрос о переименовании системы HLA в главный комплекс генов иммунного ответа человека, но, учитывая, что старое историческое название давно укоренилось среди исследователей, решено было не менять его [35].
1.1.1 Строение системы HLA
По современным представлениям система HLA, обеспечивая регуляцию иммунного ответа, осуществляет такие важнейшие физиологические функции, как взаимодействие всех иммунокомпетентных клеток организма, распознавание своих и чужеродных, в том числе измененных собственных, клеток, запуск и реализацию иммунного ответа и, в целом, обеспечивает выживание человека как вида в условиях экзогенной и эндогенной агрессии [29].
Все многообразие указанных функций обеспечивается строением главного комплекса гистосовместимости [13].
Система HLA, открытая более 40 лет назад, по-прежнему остается одной из самых сложных, наиболее хорошо изученных и вместе с тем загадочных генетических структур в геноме человека. Так, если еще в 1987 г. расстояние между его условными границами оценивалось в 2000 кб, то на сегодняшний день оно расширено более чем в 2 раза, причем протяженность отдельных его элементов - генных кластеров - колеблется в широких пределах в зависимости от HLA-гаплотипа [23].
Комплекс генов HLA (главного комплекса гистосовместимости человека) компактно расположен на коротком плече 6-й аутосомной хромосомы, занимает 3500 kb (тысяч пар оснований) и содержит более 220 генов.
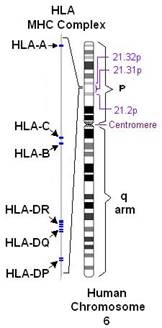
Рисунок 1. Расположение комплекса генов HLA на 6 хромосоме человека.
Гены главного комплекса гистосовместимости человека подразделяются на области A, B, С, D и обозначаются как HLA-А, HLA-В HLA-С и HLA-D, которые достаточно полиморфны и имеют варианты (аллели) и подварианты.
В последние годы в пределах I класса системы HLA открыты новые (неклассические) локусы - E, F, G, H (псевдоген, экспрессируется на уровне РНК), J (псевдоген), для большинства из которых пока не выявлено наличие аллельного полиморфизма и не определены биологические функции. Гены MIC A и MIC B расположены рядом с генами MHC I класса, но отличаются от них экзон-интронной организацией. Молекулы MIC A экспрессируются в основном на фибробластах и эпителиальных клетках. Тем не менее, предполагают, что цепь молекулы MIC A сходна с цепями молекул MHC I класса и способна связывать пептиды и другие короткие лиганды.
Между генами HLA I и II классов расположены гены, кодирующие молекулы III класса. Среди них выделяют суперсемейства C (факторы комплемента, которые вовлечены в процессы элиминации чужеродных антигенов) и G (функции окончательно не выяснены, но предполагается, что продукты экспрессии отдельных генов данного семейства участвуют в процессе созревания лейкоцитов).
На основании исследования главных комплексов гистосовместимости различных видов организмов был сделан вывод о том, что этот генный комплекс расширялся за счет дупликации, что в свою очередь давало определенные преимущества организмам с более полиморфной системой HLA в процессе эволюции. Однако вопрос о том, какие причины привели к подавлению экспрессии ранее функционировавших генов в составе МНС, остается открытым [35].
1.1.2 Характеристика генов и антигенов HLA II класса
Молекулы HLA II класса кодируются генными локусами DR, DP и DQ и имеют распространение преимущественно на макрофагах, В-лимфоцитах, активированных Т-клетках (CD4) и участвуют в гуморальном ответе. HLA-DR кодируется генами на хромосоме 6 в области 6p21.31. HLA-DR часто вовлечен в ассоциации с аутоиммунными болезнями, является протективным или ассоциированным с рядом заболеваний вместе с DQ. В случае инфекции пептид (типа энтеротоксина стафилококка) связывается с молекулой DR и DQ и представляется нескольким из очень многих TCR T-хелпера. Эти лимфоциты тогда связываются с антигенами на поверхности B-клеток, стимулируя быстрое увеличение B-клеток.
Молекулы HLA II класса - это мембранные гликопротеины, состоящие из двух цепей (α и β), каждая из которых содержит по два домена. Цепи молекул II класса очень сходны между собой. На аминокислотном уровне наиболее полиморфными являются внешние альфа-1 и бета-1 домены, которые образуют пептид-связывающую бороздку и отвечают за презентацию антигенов.
Трехмерная структура молекулы II класса была установлена в 1993 году Брауном на основе рентгеноструктурного анализа. Антигенсвязывающую бороздку молекул II класса образуют наиболее экспонированные внешние домены, α-1 и β-1. Дно бороздки сформировано b-складчатой структурой, состоящей из восьми антипараллельных участков, а стенки образованы a-спиралями (рисунок 2). Отличие антигенсвязывающей бороздки молекул II класса от таковой молекул I класса заключается в том, что в молекулах II класса она образована двумя разными цепями. Домены, образующие бороздку, и в особенности первый домен b-цепи, чрезвычайно полиморфны. Области полиморфизма собраны в несколько дискретных гипервариабельных участков (например, в b-цепи HLA-DQ они соответствуют аминокислотным остаткам в положениях 52-58, 70-77, 84-90).