Доклад: Эволюция энергетических процессов у эубактерий
Проблема акцептора электронов
Основная проблема всех процессов брожения — проблема акцептора электронов. В конечном итоге степень окисления и сопряженное с этим количество свободной энергии, а также характер образующихся продуктов определяются природой конечных акцепторов электронов. При брожениях конечными акцепторами электронов служат в основном органические соединения: метаболиты, образующиеся из исходных субстратов (пировиноградная кислота, ацетальдегид), или вещества, имеющиеся в среде культивирования (некоторые аминокислоты и другие органические соединения, способные восстанавливаться). В ряде брожений акцепторами электронов служат молекулы CO2 , а также ионы водорода (H2 ). Кроме того, в отдельных случаях дополнительными акцепторами электронов могут быть некоторые достаточно окисленные неорганические соединения, такие как нитрат, молекулярная сера. Если конечным акцептором электронов является ацетальдегид, образуется этанол, если пируват — молочная кислота. Акцептирование электронов молекулами CO2 приводит у разных видов к возникновению формиата или ацетата, если же эту функцию выполняют ионы водорода, образуется молекулярный водород (H2 ).
Восстановленные соединения, акцептировавшие электроны, выделяются из клеток эубактерий в окружающую среду и накапливаются в ней в значительных количествах. Из-за низкого энергетического выхода процессов брожения для обеспечения энергией всех функций и биосинтетических процессов клетке приходится перерабатывать огромные количества субстратов.
Итак, брожение — это способ получения энергии, при котором АТФ образуется в процессе анаэробного окисления органических субстратов в реакциях субстратного фосфорилирования.
ГОМОФЕРМЕНТАТИВНОЕ МОЛОЧНОКИСЛОЕ БРОЖЕНИЕ
Последовательность биохимических реакций, лежащих в основе гомоферментативного молочнокислого брожения, получила название гликолитического пути (гликолиза)41 , фруктозодифосфатного пути, или пути Эмбдена — Мейергофа — Парнаса (Н. Embden, О. Meyerhof, Я. О. Парнас), по именам исследователей, внесших больший вклад в изучение этого процесса. Общая схема гомоферментативного молочнокислого брожения представлена на рис. 53.
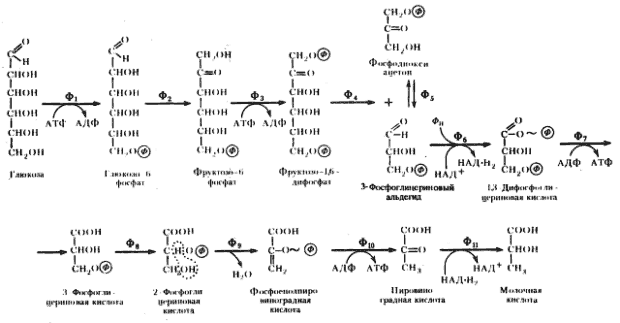 |
| Рис. 53. Гомоферментативное молочнокислое брожение: Ф1 — гексокиназа; Ф2 — глюкозофосфатизомераза; Ф3 — фофсофруктокиназа; Ф4 — фруктозо-1,6-дифосфатальдолаза; Ф5 — триозофосфатизомераза; Ф6 — 3-ФГА-дегидрогеназа; Ф7 — фофсоглицерокиназа; Ф8 — фосфоглицеромутаза; Ф9 — енолаза; Ф10 — пируваткиназа; Ф11 — лактатдегиброгеназа (по Dagley, Nicholson, 1973) |
Основными энергетическими ресурсами для эубактерий, осуществляющих гомоферментативное молочнокислое брожение, служат моносахара (в первую очередь, глюкоза) и дисахара (мальтоза, лактоза). В процессе подготовки к энергетическим преобразованиям дисахара ферментативным путем расщепляются до моносахаров. Различные моносахара, прежде чем подвергнуться преобразованиям, должны превратиться в глюкозо-6-фосфат. Момент унификации, т. е. превращения различных субстратов в один, исходный для дальнейшего его метаболизирования по данному пути, очень важен. От того, что служит исходным энергетическим ресурсом, зависит общий энергетический баланс процесса.
41 Собственно гликолиз — это определенная последовательность ферментативных реакций от углевода до пировиноградной кислоты, поэтому, строго говоря, "гликолиз" не является синонимом "гомоферментативного молочнокислого брожения", но 10 из 11 реакций у этих процессов идентичны.
Если исходным энергетическим субстратом служит глюкоза, первое превращение, которому она подвергается, — фосфорилирование. В результате образуется глюкозо-6-фосфат — метаболически активная форма глюкозы. Если исходный энергетический субстрат — лактоза, первым шагом на пути метаболизирования является ферментативное расщепление лактозы с помощью (3-галактозидазы на D -галактозу и D -глюкозу. D -галактоза затем подвергается фосфорилированию, приводящему к образованию D -галактозо-1-фосфата. Последний подвергается серии ферментативных превращений с участием УТФ в качестве кофермента, в результате которых превращается в глюкозо-1-фосфат.
У некоторых бактерий из рода Lactobacillus имеется фермент мальтозофосфорилаза, катализирующий реакцию:
мальтоза + H3 PO4 ® глюкозо-1-фосфат + глюкоза.
В результате этой реакции осуществляется расщепление дисахарида мальтозы на две молекулы глюкозы, одна из которых образуется в фосфорилированной форме. Здесь важно подчеркнуть, что в этой реакции молекула фосфорилированной глюкозы синтезируется без затраты АТФ.
Если исходным энергетическим субстратом, вовлекаемым в процесс гликолиза, служит полисахарид типа гликогена или крахмала, его использование начинается с фосфоролитического отщепления глюкозного остатка, протекающего по схеме:
(глюкоза)n + HPO4 2 – ® (глюкоза)n – 1 + глюкозо-1-фосфат.
Глюкозо-1-фосфат, образующийся в результате подготовительных превращений углеводов, иных, чем глюкоза, превращается затем в глюкозо-6-фосфат. Перемещение фосфатной группы из положения 1 в положение 6 катализируется ферментом фосфоглюкомутазой. Дальнейшее превращение глюкозо-6-фосфата одинаково независимо от исходного энергетического субстрата (рис. 53).
Молекула глюкозо-6-фосфата изомеризуется в молекулу фруктозо-6-фосфата. Последний фосфорилируется в положении 1. Донором фосфата служит АТФ. Вторичное фосфорилирование молекулы фруктозы приводит к ее дальнейшему активированию.
Образовавшийся фруктозо-1,6-дифосфат разрывается на две триозы: фосфодиоксиацетон и 3-ФГА. Разрыв катализируется фруктозо-1,6-дифосфатальдолазой (альдолазой), являющейся ключевым ферментом этого пути. Достаточно обнаружить альдолазу, чтобы получить свидетельство существования гликолитического пути у организма. В последующие реакции может включаться только 3-ФГА. Фосфодиоксиацетон превращается в 3-ФГА в реакции изомеризации, катализируемой триозофосфатизомеразой.
На этом этапе заканчивается подготовительная стадия гликолитического пути: молекула глюкозы после активирования и расщепления на 2 фосфотриозы подготовлена для последующих превращений. Для активирования 1 молекулы глюкозы тратятся 2 молекулы АТФ42 . Таким образом, до сих пор процесс протекает с затратой энергии. Однако его смысл и назначение заключаются в обеспечении клетки энергией. Эта задача решается на следующей стадии.
Окисление 3-ФГА до 1,3-дифосфоглицериновой кислоты — один из наиболее важных этапов гликолитического пути, поскольку именно на этом этапе энергия, освобождающаяся при окислении альдегидной группы 3-ФГА, запасается в молекуле 1,3-ФГК. Реакция катализируется ферментом 3-ФГА-дегидрогеназой:
3-ФГА + НАД+ + ФН ® 1,3-ФГК + НАД-H2 .
3-ФГА служит донором электронов, которые переходят на НАД+ функционирующий в качестве переносчика электронов от 3-ФГА к пировиноградной кислоте. Образование последней происходит на более поздних этапах гликолитического пути. Итак, альдегидная группа 3-ФГА окисляется до карбоксильной группы. Однако вместо свободной карбоновой кислоты образуется смешанный ангидрид фосфорной кислоты и карбоксильной группы 3-ФГК — 1,3-ФГК. Реакция окисления 3-ФГА до 1,3-ФГК с помощью НАД-зависимой 3-ФГА-дегидрогеназы состоит из нескольких этапов, в результате чего энергия, освобождающаяся при окислении 3-ФГА, запасается в макроэргической фосфатной связи у первого углеродного атома 1,3-ФГК. 1,3-ФГК реагирует далее с АДФ, отдавая высокоэнергетическую фосфатную группу, что приводит к синтезу молекулы АТФ. Таким образом, энергия, высвободившаяся при окислении альдегидной группы, оказывается запасенной в молекуле АТФ.
42 Если исходным субстратом служит полисахарид, например гликоген или крахмал, для активирования глюкозного остатка на подготовительной стадии гликолитического пути затрачивается только 1 молекула АТФ.
Итак, произошло образование 3-ФГК. Теперь можно подвести некоторые итоги. Клетка на этом этапе "вернула" свои энергетические затраты: 2 молекулы АТФ были затрачены и 2 молекулы АТФ синтезировались на 1 молекулу глюкозы. На этом же этапе в реакции окисления 3-ФГА до 1,3-ФГК и образования АТФ имеет место первое субстратное фосфорилирование. Энергия освобождается и запасается в макроэргических фосфатных связях АТФ в процессе перестройки сбраживаемого субстрата при участии ферментов. Реакция, ведущая к субстратному фосфорилированию, может быть проведена в пробирке. Все необходимые для этого компоненты известны и получены в чистом виде. Возможность осуществления реакции в пробирке указывает на то, что фермент, катализирующий ее, не связан с клеточными структурами. Первое субстратное фосфорилирование носит еще название фосфорилирования на уровне 3-ФГА.
После образования 3-ФГК фосфатная группа из третьего положения переносится во второе. Далее происходит отщепление молекулы воды от второго и третьего атомов углерода 2-ФГК, катализируемое ферментом енолазой, и образуется фосфоенолпировиноградная кислота. В результате происшедшей дегидратации молекулы 2-ФГК степень окисления ее второго углеродного атома увеличивается, а третьего — уменьшается. Таким образом, данная реакция по существу представляет собой внутримолекулярный окислительно-восстановительный процесс. Дегидратация молекулы 2-ФГК, приводящая к образованию ФЕП, сопровождается перераспределением энергии внутри молекулы, в результате чего фосфатная связь у второго углеродного атома из низкоэнергетической в молекуле 2-ФГК превращается в высокоэнергетическую в молекуле ФЕП.
Молекула ФЕП становится донором богатой энергией фосфатной группы, которая переносится на АДФ с помощью фермента пируваткиназы. Таким образом, в процессе превращения 2-ФГК в пировиноградную кислоту имеет место высвобождение энергии и запасание ее в молекуле АТФ. Это второе субстратное фосфорилирование. По ряду черт оно отличается от первого субстратного фосфорилирования: 1) если в первом случае образование макроэргической фосфатной связи протекало одновременно с присоединением к субстрату фосфатной группы, то во втором — фосфатная группа была присоединена к молекуле субстрата задолго до этого события; 2) первое субстратное фосфорилирование связано с реакцией окисления, приводящей к тому, что от молекулы 3-ФГА отрываются два электрона и переходят на НАД+ , т. е. молекула 3-ФГА служит донором электронов, но вопрос о конечном акцепторе их на этом этапе не решен. Напротив, при втором субстратном фосфорилировании, связанном с реакцией дегидратации молекулы 2-ФГК, решается проблема и донора и акцептора. Здесь в результате внутримолекулярного окислительно-восстановительного процесса одна молекула и донирует и акцептирует электроны.
В процессе второго субстратного фосфорилирования образуется еще молекула АТФ; в итоге общий энергетический выигрыш процесса составляет 2 молекулы АТФ на 1 молекулу глюкозы. Такова энергетическая сторона процесса гомоферментативного молочнокислого брожения.
Однако осталась еще проблема восстановленного переносчика — НАД-H2 , образованного в реакции окисления 3-ФГА. Чтобы процесс продолжался, в метаболический поток необходимо вернуть этот метаболит в окисленном виде (НАД+ ), т. е. решить проблему конечного акцептора. Как же она решается в данном случае? Результатом рассмотренного выше процесса, помимо его энергетического итога, является образование 2 молекул пировиноградной кислоты и 2 молекул НАД-H2 на 1 молекулу сброженной гексозы. Молекула пировиноградной кислоты по своему химическому строению — достаточно окисленное соединение и может служить акцептором электронов. В этом случае донор-акцепторная проблема решена самым простым способом: 2 электрона переносятся с НАД-H2 на молекулу пировиноградной кислоты, что приводит к образованию молочной кислоты. Суммарно процесс можно выразить в виде следующего уравнения:
глюкоза + 2ФН + 2АДФ ® 2 молочная кислота + 2АТФ + 2H2 O.
Гомоферментативное молочнокислое брожение представляет собой энергетическую сторону образа жизни группы гомоферментативных молочнокислых бактерий. Черты древности этой группы видны не только в процессе добывания ее представителями энергии, но и в других сторонах их метаболизма, о чем будет сказано в разделе, посвященном краткой характеристике этих бактерий. Сейчас же остается подвести некоторые итоги рассмотренного процесса и оценить его "судьбу". В процессе гомоферментативного молочнокислого брожения имеют место 3 типа химических превращений:
- перестройка углеродного скелета исходного субстрата;
- окислительно-восстановительные превращения;
- образование АТФ.