Доклад: Неканонические наследственные изменения
Приобретенные признаки наследуются
“История биологии не знает более выразительного примера многовекового обсуждения проблемы, чем дискуссия о наследовании или о ненаследовании приобретенных признаков”, - эти слова стоят в начале книги известного цитолога и историка биологии Л.Я.Бляхера [12]. В истории, пожалуй, можно вспомнить аналогичную ситуацию с попытками превращения химических элементов. Алхимики верили в эту возможность, но в химии утвердился постулат о неизменности химических элементов. Однако ныне в ядерной физике и химии исследования по превращению элементов и анализ их эволюции - дело обычное. Кто же оказался прав в многовековом споре? Можно сказать, что на уровне химических молекулярных взаимодействий не происходит превращения элементов, а на ядерном уровне оно - правило.
Напрашивается сходная аналогия и с вопросом о наследовании признаков, которые появились в ходе онтогенеза. Если вновь возникающие наследственные изменения сводить лишь к мутациям генов и хромосом, тогда вопрос можно считать закрытым. Но если исходить из обобщенной концепции генома, включая представление о динамической наследственности [2, 3], проблема нуждается в пересмотре. Помимо мутационной существует вариационная и эпигенетическая формы наследственной изменчивости, связанные не с изменениями в тексте ДНК, а в состоянии гена. Такие эффекты обратимы и наследуемы.
Интересно, что вышедший в конце 1991 г. Международный ежегодник по генетике открывается статьей О.Ландмана “Наследование приобретенных признаков” [13]. Автор суммирует уже давно полученные в генетике факты, показывая, что “наследование приобретенных признаков вполне совместимо с современной концепцией молекулярной генетики”. Ландман детально рассматривает около десяти экспериментальных систем, в которых установлено наследование приобретенных признаков. Четыре разных механизма способны привести к нему: изменение структур клеточной оболочки, или кортекса, изученное Т. Соннеборном у инфузорий; ДНК-модификации, т.е. клонально передаваемые изменения в характере локального метилирования ДНК (сюда входит феномен импринтинга); эпигенетические изменения без каких-либо модификаций ДНК; индуцированная утрата либо приобретение факультативных элементов.
Статья Ландмана делает нас как бы свидетелями критического периода смены постулата в генетике, казавшегося неколебимым как скала. Автор спокойно, без ажиотажа и новых сногсшибательных фактов, объединяет старые и новые данные в систему, дает им ясное современное истолкование. Можно сформулировать общий принцип: наследование приобретенных признаков возможно в тех случаях, когда некий фенотипический признак зависит от числа или топографии факультативных элементов.
Приведу два поучительных примера на дрозофиле: первый связан с поведением вируса сигма, второй - мобильных элементов, ответственных за гибридную стерильность самок и сверхмутабильность.
Изучение взаимодействия вируса сигма с геномом дрозофилы началось более 60 лет назад. Сначала в 1937 г. французский генетик Ф.Леритье обнаружил резкие наследственные отличия у разных линий мух по степени чувствительности к углекислому газу (СО2 ). Признак наследовался причудливым образом: через цитоплазму, но не только по материнской линии, а иногда и через самцов. Чувствительность можно было передать и путем инъекции гемолимфы, причем разным видам дрозофил. В этих случаях признак передавался не стабильно, но в результате отбора наследование становилось устойчивым.
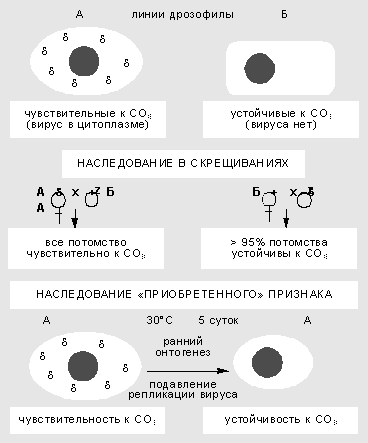
Неменделевское наследование признака у дрозофилы, который зависит от популяции факультативных элементов генома. Признак чувствительности к СО2 вызван присутствием в цитоплазме мухи рабдовируса сигма. В результате температурного шока на ранней стадии развития дрозофилы блокируется размножение вируса, и выросшие особи приобретают устойчивость к нему.
Чувствительность к СО2 оказалась связана с устойчивым размножением в половых и соматических клетках РНК-содержащего пулевидного рабдовируса сигма, сходного по ряду свойств с вирусом бешенства у млекопитающих. Оогонии (клетки, из которых в ходе мейоза и созревания образуются яйцеклетки) у самок стабилизированной линии обычно содержат 10-40 вирусных частиц, а ооциты (зрелые яйцеклетки) - 1-10 млн. Вирус сигма - типичный факультативный элемент. Мутации в его геноме приводят к сложным формам поведения системы. Найдены случаи вирусоносительства, при которых дрозофилы остаются устойчивы к СО2 , но вместе с тем иммунны к заражению другими штаммами вируса. Ситуация вполне сравнима с поведением системы фаг-бактерия, что сразу заметили Ф.Жакоб и Э.Вольман.
Взаимоотношения генома дрозофилы и размножающегося в ее цитоплазме вируса подчиняются правилам внутриклеточной генетики. Воздействия в ходе онтогенеза могут вызвать сдвиг в числе и межклеточной топографии частиц и как результат - изменить степень чувствительности к углекислому газу. Так, повышенная температура блокирует репликацию вирусных частиц. Если самок и самцов в период гаметогенеза содержать несколько дней при температуре 30°С, потомство от таких мух будет свободно от вируса и устойчиво к СО2 . Значит, приобретенный в ходе индивидуального развития признак наследуется в ряду поколений.
Ситуация с вирусом сигма не единична. Французские генетики изучали факторы стерильности самок, связанные с поведением мобильных элементов типа “I”. Наследование этого признака определяется сложными ядерно-цитоплазматическими взаимодействиями. Если в отцовских хромосомах локализованы активные I-элементы, то на фоне R-цитоплазмы они начинают активироваться, претерпевают множественные транспозиции и в результате вызывают резкие нарушения онтогенеза в потомстве самок с чувствительной цитоплазмой. Такие самки откладывают яйца, но часть эмбрионов гибнет на ранней стадии дробления - еще до образования бластомеры. Линии, выделенные из природных популяций, отличаются по силе действия I-факторов и степени реактивности (или чувствительности) цитоплазмы. Эти показатели можно изменить внешним влиянием. Возраст исходных родительских самок, а также воздействие в ранний период развития повышенной температуры отражаются не только на плодовитости выросших самок, но и на плодовитости их потомства. Вызванные условиями среды изменения реактивности цитоплазмы поддерживаются на протяжении многих клеточных поколений. “Самое замечательное, что эти изменения реактивности цитоплазмы под влиянием негенетических факторов наследуются: наблюдается наследование “благоприобретенных” признаков”, - отмечал Р.Б.Хесин [5].
Наследование через цитоплазму: от бабушек до внуков
В теории развития и феногенетике ХХ в. важное место занимают глубокие и совершенно оригинальные исследования эмбриолога П.Г.Светлова (1892-1972). Остановимся на разработанной им теории квантованности онтогенеза (наличии критических периодов в развитии, когда происходит детерминация морфогенетических процессов и одновременно повышается чувствительность клеток к повреждающим агентам) и на развитой в связи с этим идее, что изучение онтогенеза надо вести не с момента оплодотворения и образования зиготы, а еще с гаметогенеза, включающего оогенез у самок предшествующего поколения - проэмбрионального периода.
На основании этих постулатов Светлов провел в 60-е годы простые и ясные опыты на дрозофиле и мышах. Он убедительно показал, что возможно стойкое неменделевское наследование свойств цитоплазмы, а модификации в выраженности мутантных признаков, возникшие после кратковременного внешнего воздействия в критический период развития организма, тоже передаются в ряду поколений [14].
В одной из серий опытов он сравнивал степень проявления мутантного признака в потомстве двух линий мышей, гетерозиготных по рецессивной мутации микрофтальмии (уменьшенный размер сетчатки и глаз с момента рождения): нормальных по фенотипу гетерозигот, у которых мутантными были матери, и тех, у которых мутантны отцы. Потомство от мутантной бабушки отличалось более сильным проявлением признака. Светлов объяснял этот странный факт тем, что женские гаметы гетерозиготных самок находились еще в теле их мутантных матерей и испытывали с их стороны влияние, которое усилило мутации у внуков.
По существу Светлов установил явление, впоследствии получившее название “геномный импринтинг” - различие в выраженности гена в зависимости от того, пришел он к потомству от матери или от отца. Работы эти, увы, остались недооцененными.
Интересно, что еще в конце 80-х годов импринтинг, как остроумно заметил К.Сапиенца, исследователь этого феномена, было “принято считать генетическим курьезом, затрагивающим лишь очень немногие признаки. Меня неоднократно спрашивали, почему я попросту трачу свое время на столь незначительное явление” [15]. Большинство исследователей безоговорочно принимали одно из главных положений Менделя - “зачаток”, или ген, не может менять свои потенции в зависимости от пола, на чем основано повсеместно наблюдаемое расщепление 3:1. Но Сапиенца совершенно справедливо заметил, что при анализе менделевского расщепления обычно рассматривают только наличие ил?