Курсовая работа: Біологія та екологія бактеріофагів
Основним компонентом вірусів є також нуклеїнові кислоти (РНК або ДНК). Раніше вважали, що віруси рослин містять РНК. Однак у 1968 р. було виявлено, що вірус мозаїки цвітної капусти містить дволанцюгову молекулу ДНК. Більшість вірусів рослин містять одноланцюгову лінійну РНК. В одних видів тваринних вірусів знайдено ДНК, в інших — РНК. Бактеріофаги містять ДНК.
На відміну від клітинних організмів, у вірусів, на додаток до двох основних типів нуклеїнових кислот, є також одноланцюгові ДНК і дволанцюгові РНК.
Одноланцюгова ДНК фагів має кільцеву форму, а вірусів, які уражають еукаріотні клітини, — лінійну. Вторинна структура одно-ланцюгової ДНК вірусного походження нічим не відрізняється від макромолекулярної організації одноланцюгових РНК.
Дволанцюгова РНК, вперше відкрита у реовірусів, виявилась досить поширеною в царстві вірусів. Вторинна структура РНК являє собою досконалу подвійну спіраль, що містить два комплементарні полірибонуклеотидні ланцюги.
Слід зазначити, що дволанцюгова ДНК Т-парних фагів відрізняється від ДНК інших вірусів тим, що замість цитозину містить 5-оксиметилцитозин і глюкозу, приєднану до деяких оксиметильних груп оксиметилцитозину.
Крім білків і нуклеїнових кислот, до складу вірусних частинок можуть входити ліпіди, вуглеводи та інші речовини. Ліпіди виявлені у складно організованих вірусів. Вони містяться переважно в складі ліпопротеїдної оболонки (суперкапсида) і формують її ліпідний бімолекулярний шар, в якому містяться суперкапсидні білки.
Вважають, що ліпідний компонент стабілізує структуру вібріону. Характерним для вірусів, які містять ліпіди, є те, що без винятку всі ліпіди у них не кодуються геномом, а мають клітинне походження.
Вуглеводи у вірусів входять до складу глюкопротеїдів. Кількість їх може становити понад 10 % у перерахунку на масу вібріону.
Вуглеводний компонент глюкопротеїдів вірусу відіграє істотну роль у структурі та функції білка. Він є каркасом для локальних ділянок глюкопротеїду, забезпечуючи збереження конформації білкової молекули, і зумовлює її захист від ферментів протеаз.
Віруси — облігатні внутрішньоклітинні паразити, тобто вони можуть рости і розвиватися тільки всередині живих клітин хазяїна. Ось чому протягом багатьох років, починаючи з досліджень Д. Й. Івановського, вченим не вдавалося виростити віруси на звичайних природних і синтетичних поживних середовищах, на яких добре культивуються бактерії, гриби та інші мікроорганізми. Деякі віруси людини ще й досі не вдається культивувати в лабораторних умовах.
З метою вирощування і нагромадження вірусів тварин, рослин і мікроорганізмів широко застосовуються культури органів, клітин, ембріони курчат, лабораторні тварини, сприйнятливі рослини тощо.
1.3 Репродукція вірусів
Зважаючи на складність процесу репродукції та його кардинальну відмінність від розмноження прокаріотів, вчені розробили низку методів для вивчення проникнення вібріонів у живу клітину хазяїна. Часто використовують метод змішування певної кількості вірусів із суспензією клітин з наступним відокремленням клітин центрифугуванням і визначенням вмісту вірусів у надосадовій рідині. При використанні хімічних методів застосовують так званий мічений вірус, в якого один з компонентів (білок або нуклеїнова кислота) має радіоактивну мітку. З морфологічних методів найчастіше використовують електронну мікроскопію.
Завдяки сучасним методам дослідження вченим вдалося встановити складний характер взаємодії вірусу з клітиною хазяїна. Залежно від властивостей вірусу і сприйнятливої клітини, а також умов зовнішнього середовища, розрізняють такі основні типи взаємодії вірусу з клітиною: а) розмноження вірусу призводить до руйнування і загибелі клітини (такий тип взаємодії дістав назву продуктивної інфекції, а віруси — вірулентних); б) взаємодія, при якій клітини залишаються живими, а дозрілі вібріони не утворюються, називається абортивною інфекцією; в) геном вірусу інтегрується (об'єднується) з генетичним апаратом клітини і під час поділу клітини може тривалий час передаватися в дочірні клітини. Однак через деякий час, за певних умов, у клітині може початися розмноження вірусу, яке призводить клітину до загибелі. Такий тип взаємодії дістав назву вірогенії.
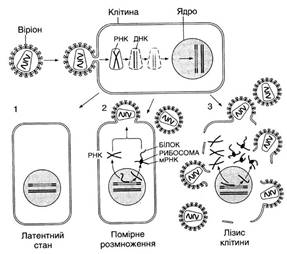
Рис. 4. Схема розмноження вірусу СНІД (за У.А. Хезелтайн, Ф. Вонг-Сталь, 1988):
1 — заражена клітина перебуває в латентному стані; 2 - у клітині починається помірне розмноження вірусу; 3 — лізис (руйнування) клітини і вихід вірусних частинок назовні. Інфекція ВІЛ починається з того, що вібріон приєднується до сприйнятливої клітини. При цьому оболонка вірусу і мембрана клітини зливаються так, що серцевина вірусної частинки, яка містить крім білків генетичний матеріал вірусу — РНК, опиняється в цитоплазмі клітини. Тут зворотна транскриптаза, яка міститься в серцевині, синтезує одноланцюгову ДНК, комплементарну вірусній РНК. Далі ця РНК розщеплюється і синтезується другий ланцюг, що є комплементарним першому. Дволанцюгова вірусна ДНК (провірус) проникає в ядро і вбудовується в хромосомну ДНК клітини. В такій латентній (прихованій) формі вірус може існувати, не виявляючи себе, тривалий час (1). У випадку активації провірусу, за допомогою клітинного механізму транскрипції утворюються РНК-копії провірусу і мРНК. За цими мРНК на рибосомах клітини синтезуються вірусні білки, з них і РНК-копій формуються нові віріони. Вони покидають клітину, відбруньковуючись від її поверхні. Якщо репродукція ВІЛ відбувається помірно, то заражена клітина може залишатися цілою і життєздатною (2). Якщо ж реплікація вірусу буде дуже інтенсивною, то відбувається руйнування (лізис) клітини, і вона гине (3)
При першому типі взаємодії розмноження вірусу відбувається шляхом послідовного перебігу таких стадій: 1) адсорбція вірусу на клітинах; 2) проникнення вірусу в клітину; 3) роздягання (депротеїнізація оболонки) вірусу в клітині; 4) транскрипція нуклеїнової кислоти; 5) трансляція інформаційних РНК; 6) реплікація генома; 7) складання вірусних компонентів і вихід дозрілих вібріонів з клітини хазяїна (рис. 4).
Адсорбція. Першу стадію проникнення вірусів у клітину — прикріплення їх до клітинної стінки, можливе через електростатичну взаємодію позитивно і негативно заряджених угруповань на поверхні вірусу і клітини, — називають адсорбцією. Це високо специфічний процес. Структури вібріону, що уможливлюють його адсорбцію на клітині, називаються вірусними рецепторами або прикріплюючими білками, оскільки вони «впізнають» специфічні угруповання на мембрані клітини і зумовлюють прикріплення до них вірусу.
Кількість специфічних рецепторів на поверхні клітини може складати до 105 на одну клітину. Наявність відповідних рецепторів — один із важливих факторів, що визначають можливість чи неможливість для даного вірусу спричиняти інфекційний процес у клітині. Саме відсутністю специфічних рецепторів пояснюють стійкість деяких клітин і організмів до зараження певними вірусами.
Після того, як вібріон адсорбувався на поверхні клітини, починається друга стадія — проникнення його в клітину. Перехід цілого вібріону крізь цитоплазматичну мембрану може відбуватися шляхом віропексису (піноцитозу) або внаслідок злиття цитоплазматичної мембрани з оболонкою вібріону. Вважається, що ці механізми доповнюють один одного. Фітопатогенні віруси можуть проникати в рослинну клітину за допомогою комах-переносників при вегетативному розмноженні, під час догляду за рослинами тощо, а деякі бактеріофаги проникають крізь клітинну стінку шляхом ін'єкції.
Більшість оболонкових і без оболонкових вірусів тварин проникають у клітину шляхом рецепторного ендоцитозу, який відбувається в певних ділянках цитоплазматичної мембрани, в спеціальних заглибленнях, на дні яких містяться специфічні рецептори. Заглиблення забезпечують швидку інвагінацію і утворення вкритих особливим білком, клатрином, внутрішньоклітинних вакуолей. Ендоцитоз уможливлює внутрішньоклітинний транспорт вірусу в складі ендоцитарної вакуолі. Остання може рухатися в будь-якому напрямі і зливатися з клітинними мембранами, і при цьому звільнювати вірусну частинку у відповідних ділянках всередині клітини.
Депротеїнізація оболонки. Наступною стадією є «роздягання», тобто звільнення нуклеїнової кислоти вірусу від захисної оболонки, яка перешкоджає експресії вірусного геному. «Роздягання» низки вірусів відбувається за допомогою ферментів у спеціалізованих органелах клітини (лізосомах, диктіосомах), перинуклеарному просторі, ядерних порах, на ядерній мембрані тощо. Цей поступовий процес є результатом серії послідовних реакцій.
Транскрипція. У наступній стадії здійснюється процес транскрипції звільненої від оболонок нуклеїнової кислоти вірусу, тобто переписування інформації, яка міститься в ДНК, на РНК за законами генетичного коду і за допомогою РНК-полімерази. Продуктами транскрипції в клітині є іРНК. У ДНК-вмісних вірусів іРНК синтезується на матриці однієї із ниток ДНК. Схема перенесення генетичної інформації у цих вірусів така ж, як і в клітини:
![]()
РНК-вмісні віруси, у яких генетична інформація зберігається не в ДНК, а в РНК, зокрема у РНК-вмісних «плюс-ниткових» вірусів, що в них функції іРНК виконує сам геном, генетична інформація передається за такою схемою: РНК → білок (пікорнавіруси, тогавіруси, коронавіруси). У вірусів, геном яких не може виконувати функцію іРНК, у клітині синтезується комплементарна геному іРНК. У цьому випадку передача генетичної інформації здійснюється у такий спосіб: РНК → іРНК → білок. Сюди належать ортоміксовіруси, пара-міксовіруси та ті віруси, у яких геном представлений одноланцюговою РНК, а також реовіруси та деякі інші, геном яких складається з дволанцюгової РНК. Деякі віруси тварин — так звані ретровіруси — мають унікальний шлях передачі генетичної інформації. У них РНК переписується на ДНК, а ДНК інтегрує з геномом клітини і в його складі переписується на РНК, яка володіє функціями іРНК. Такі віруси мають вірусоспецифічний фермент, який переписує РНК на ДНК. Цей процес одержав назву зворотної транскрипції, а фермент — зворотної транскриптази (ревертаза). Вона також може синтезувати нитку ДНК на матриці ДНК (рис. 4).
Транскрипція деяких ДНК-вмісних вірусів (аденовірусів, вірусів герпесу та інших) відбувається в ядрі клітини за допомогою клітинних ферментів РНК-полімерази II та РНК-полімерази III. У РНК-вмісних вірусів транскрипція здійснюється вірусоспецифічними транскриптазами, які закодовані у вірусному геномі. Транскрипція вірусного генома регулюється протягом інфекційного циклу як клітинними, так і вірусоспецифічними механізмами.
Трансляція. Процес переведення генетичної інформації, яка міститься в іРНК, у специфічну послідовність амінокислот у молекулі білка називається трансляцією. Саме в результаті трансляції іРНК відбувається синтез білка в клітині. Він здійснюється на рибосомі, яка складається з двох субодиниць — великої і малої. Субодиниці містять рибосомальні РНК і білки.
У клітині завжди є РНК, які транспортують в рибосоми амінокислоти, їх називають транспортними (тРНК). Кожна амінокислота переноситься відповідною тРНК. Три нуклеотиди на іРНК, що кодують одну амінокислоту, називаються триплетом або «кодоном». На кінці тРНК є три нуклеотиди, комплементарні кодону, — це «антикодон». Останній відповідає за прикріплення до певного місця іРНК (кодону). Тільки після утворення на рибосомі потрійного комплексу: ІРНК — тРНК — АК може початися процес трансляції.
Трансляція перебігає у три фази: ініціація, елонгація і термінація.