Курсовая работа: Біологія та екологія бактеріофагів
Термінація (закінчення) трансляції здійснюється тоді, коли рибосома доходить до термінуючого кодону в складі ІРНК. У цей момент синтез поліпептидного ланцюга (білкової молекули) закінчується і він звільняється від полірибосоми. Після цього полірибосоми розпадаються на субодиниці. Останні можуть входити до складу нових полірибосом.
Реплікація (від лат. replicatio — подвоєння) — синтез молекул нуклеїнової кислоти, які гомологічні геному. Реплікація геному ДНК-вмісних вірусів каталізується переважно клітинними ферментами; її механізм подібний до реплікації клітинної ДНК. Кожна синтезована молекула ДНК складається з однієї батьківської і однієї заново синтезованої дочірньої нитки. Це так званий напівконсервативний механізм реплікації.
Реплікація одноланцюгових ДНК (у парвовірусів) проходить з утворенням дволанцюгових проміжних реплікативних форм. Останні реплікуються за класичним напівконсервативним механізмом. Найкраще вивченим з цієї групи вірусів є фаг фх 174.
У вірусів, які містять кільцеві дволанцюгові ДНК (паповавіруси), розрізується один із ланцюгів ДНК, що приводить до розкручування супервитків на певній ділянці молекули.
Реплікація вірусних РНК відбувається в клітині за допомогою вірусоспецифічних ферментів. Реплікацію каталізує той самий фермент, що й транскрипцію (репліказа). При реплікації одноланцюгових РНК спочатку синтезуються комплементарні геному ланцюги, які, своєю чергою, стають матрицями для синтезу копій генома. Механізм реплікації дволанцюгової РНК добре вивчено у реовірусів, віріони яких також містять вірусоспецифічну РНК-полімеразу. Після проникнення віріону в клітину РНК-полімераза частково звільняється від білків, активується і здійснює синтез ІРНК, причому зчитування відбувається тільки з одного ланцюга батьківських молекул РНК, тобто за консервативним механізмом.
Синтез дволанцюгових дочірніх молекул РНК здійснюється асинхронно — на матриці батьківської РНК синтезуються комплементарні її ланцюги, які є матрицями для синтезу другого ланцюга РНК.
Синтез компонентів вірусних частинок може відбуватися в різних структурах ядра і цитоплазми. У деяких вірусів синтез вірусних нуклеїнових кислот проходить в ядрі, а синтез вірусних білків — в цитоплазмі. Отже, на відміну від бактерій та інших клітинних організмів, вірусам властивий диз'юнктивний (роз'єднаний) спосіб розмноження, який полягає в тому, що їхні компоненти синтезуються в клітині хазяїна відокремлено, неодночасно в різних місцях.
Складання вібріонів і вихід їх з клітини. Завершальним етапом внутрішньоклітинного існування вірусів є формування з новосинтезованих компонентів (НК і білків) нових вібріонів з властивою їм структурою та вихід дозрілих вірусних частинок з клітини.
Упродовж еволюції у вірусів виникли два способи перебігу цього завершального етапу. В аденовірусів, пікорнавірусів, реовірусів та деяких інших складання та дозрівання вібріонів йде усередині клітини. У вірусів, які мають оболонку (РНК-вмісні віруси з негативним геномом, ретровіруси та інші), складання вібріону і вихід його з клітини відбувається шляхом відбрунькування в зовнішнє середовище. Вважається, що це найбільш ефективний механізм виходу вірусу.
При диз'юнктивному розмноженні утворення вібріону можливе лише в тому випадку, якщо вірусні НК і білки володіють здатністю при достатній концентрації «упізнавати» один одного серед різноманітних клітинних білків і НК та спонтанно з'єднуватися. Білково-нуклеїнове «упізнавання» обмежено невеликою ділянкою нуклеїнової кислоти і визначається унікальною послідовністю нуклеотидів у некодуючій частині вірусного генома.
У простих вірусів спочатку формуються провібріони, які далі, в результаті модифікації білків, перетворюються на вібріони. Найпростіший механізм складання вібріонів спостерігається у вірусів зі спіральною симетрією. Так, у класичного представника таких вірусів — ВТМ — вібріони формуються самоскладанням, тобто спонтанною агрегацією за типом кристалізації.
У складних вірусів складання вібріонів здійснюється у кілька етапів з участю ядерних і цитоплазматичних мембран. Взаємодія нуклеїнової кислоти з внутрішніми білками приводить до формування нуклеокапсидів, які є проміжними структурами при складанні. У деяких вірусів є низка гідрофобних білків, які виконують функції посередників між сформованими нуклеокапсидами і вірусними оболонками. Механізм складання вібріонів РНК- і ДНК-вмісних вірусів досить складний і досі ще недостатньо вивчений.
Останньою стадією репродукції вірусів є вихід повністю сформованих вібріонів з клітини назовні. Це відбувається шляхом «вибуху» або брунькування. При першому способі спостерігається лізис (руйнування) і загибель клітини і вихід з неї дозрілих вібріонів. Це характерно для вірусів, які не мають ліпопротеїдної оболонки (адено-, пікорна-, рео-, парвовіруси та ін.).
Вихід із клітини шляхом брунькування властивий вірусам, які мають ліпопротеїдну мембрану, що є похідною клітинних мембран. При цьому способі клітина хазяїна може тривалий час зберігати життєздатність і продукувати вірус до повного виснаження своїх ресурсів.
Розглянутий нами процес розмноження властивий переважній більшості вірулентних вірусів, основною рисою яких є здатність автономно розмножуватися в інфікованій клітині, що, як правило, призводить до загибелі клітин хазяїна. Однак існує велика група так званих помірних вірусів, характерною рисою яких є здатність до інтеграції (об'єднання) вірусного генома з геномом клітини хазяїна. В цьому разі вірус у клітині нічим не виявляє своєї присутності і тривалий час існує «приховано» (екліпс), стаючи нібито складовою частиною генетичного матеріалу клітини.
Розділ 2. Поняття про бактеріофаги, їх особливості
2.1 Морфологія фагів
Терміни “бактеріофаги” і “бактеріофагія” стали загальновизнаними. Поряд з ними в літературі широко застосовується зручний термін “фаг”, на позначення і бактеріофагів, що вражають бактерій, і для відкритих дещо пізніше актинофагів (які вражають актиноміцети), альгофагів (що вражають деякі водорості).
Припущення, що бактеріофаги мають корпускулярну природу, було висунуто ще Ф.д'Ерелєм. Однак тільки після винайдення електронного мікроскопа вдалося побачити і вивчити ультраструктуру фагів. Нагадаємо, що довгий час уявлення про морфологію та основні особливості фагів ґрунтувалися на результатах вивчення фагів Т-групи — ТІ, Т2,..., Т7, які розмножуються на Е.соlі штаму В. Однак з кожним роком з'являлися нові дані щодо морфології і структури різноманітних фагів, що зумовило необхідність їхньої морфологічної класифікації.
Детальні електронно-мікроскопічні дослідження, в поєднанні з деякими фізико-хімічними методами вивчення фагів Т-групи, показали, що кожен фаг складається з різних морфологічних елементів.
Основні частини найкраще вивчених булавоподібних фагів становлять головка з білковою оболонкою — капсидом і відросток. Субодиниці капсиду називають капсомерами. Структурні елементи складних відростків дістали назву зовнішнього чохла, внутрішнього стрижня і базальної пластинки, відростка з зубцями і нитками.
А.С. Тихоненко (1972) поділяє фаги з огляду на ускладнення їхньої структури (що з еволюційної точки зору є найбільш доцільним) на п'ять основних груп. (рис. 6)
До першої групи слід віднести ниткоподібні фаги fd, fl, M13 та ін. За формою вони нагадують вібріони ВТМ. Це довгі гнучкі палички (700—850 нм), які складаються з трубкоподібного капсиду зі спіральним типом симетрії і містять одноланцюгову ДНК.
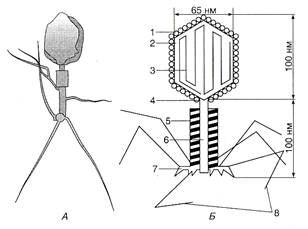
Рис 5. Структура бактеріофага Т2:
А — електронна фотографія фага Т2, Б — схема структури: 1 — білкові субодиниці капсиду; 2 — головка фага; 3 — ДНК; 4 — відросток; 5 — футляр, 6 — стрижень; ?— пластинка з шістьма зубцями, 8 — нитки виростка
Другу групу складають дрібні сферичні фаги ікосаедричної форми без диференційованого відростка. Серед них розрізняють дві підгрупи. Фаги першої підгрупи (S13, φ x 174 та ін.) мають одноланцюгову ДНК, а фаги другої підгрупи — f2, fr, MS2, R17, М12 — РНК.
До третьої групи відносять фагів з чітко вираженим хвостовим відростком невеличкого розміру. Вони інфікують бактерії, актиноміцети, хлорелу та інші організми. За будовою їхнього відростка виділяють дві групи. Представники першої групи (фаги ТЗ і Т7) мають короткий конусоподібний відросток без базальної пластинки, а представники другої (наприклад, фаг Р22 Salm. typhimurium) — короткий відросток з базальною пластинкою.
До четвертої групи належать булавоподібні фаги з довгим відростком, що не скорочується і нагадує гофровану трубку (фаги ТІ, Т5, λ та інші). Вони містять дволанцюгову ДНК.
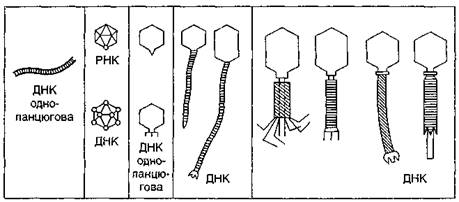
Рис. 6. Схематичне зображення представників різних груп фагів (за А.С.Тихоненком, 1968)