Курсовая работа: ЯМР-спектроскопия нуклеиновых кислот, полисахаридов и липидов
Наряду с протеинами существуют и другие макромолекулы, которые выполняют важные биологические функции. Большинство методов ЯМР, используемых для исследования протеинов, могут быть при этом непосредственно перенесены на другие макромолекулы. В этой работе рассмотрим характерные особенности применения метода ЯМР для исследования нуклеиновых кислот, полисахаридов и липидов.
1. Состав и структура нуклеиновых кислот
Нуклеиновые кислоты занимают ключевую позицию в процессах хранения и передачи информации о наследственных свойствах организма. Как и протеины, они являются линейными макромолекулами, состоящими из небольшого числа различных фрагментов, нуклеотидов.


ЯМР нуклеиновых кислот в принципе сталкивается с теми же проблемами, что и ЯМР протеинов. В то же время определение пространственной структуры нуклеиновых кислот является более простой задачей, так как здесь выбор основной пары нуклеотидов существенно ограничивает число возможных пространственных структур.
Основными фрагментами дезоксирибонуклеиновых кислот являются дезоксирибонуклеотиды, а основными фрагментами рибонуклеиновых кислот – рибонуклеотиды. По аналогии с аминокислотами в протеинах эти фрагменты отличаются только своими боковыми цепями, которые в ДНК в основном состоят из пиримидиновых производных – цитозина и тимина – и пуриновых производных – аденина и гуанина. В РНК присутствуют те же боковые цепи, только основание тимин заменено на урацил. Кроме этих основных фрагментов нуклеиновых кислот имеется еще небольшое количество производных этих оснований – так называемые редкие основания.

ДНК представляет собой двойную спираль, образованную двумя переплетающимися нитями. Она может существовать в нескольких различных формах, значение которых для процессов регуляции и контроля передачи наследственной информации в дальнейшем будет подробно обсуждаться. РНК, как правило, существует в виде мономера. Основными формами РНК являются матричная РНК, транспортная РНК и рибосомная РНК. Как ДНК, так и РНК вступаете разнообразные взаимодействия с протеинами, в результате чего происходит либо считывание информации, либо ее регуляция. Дополнительно ДНК в эукариотических клетках образуют ассоциаты с протеинами: гистонами, которые участвуют в строительстве хромосом. Подобные взаимодействия демонстрируют РНК, например рРНК связывается с протеинами, из которых состоят рибосомы. В процессах расшифровки и передачи информации мРНК и тРНК вступают в разнообразные взаимодействия с ферментами, участвующими в этих процессах. Поэтому ясно, что исследование взаимодействий между протеинами и нуклеиновыми кислотами является одной из самых интересных областей применения метода ЯМР к биологическим задачам.
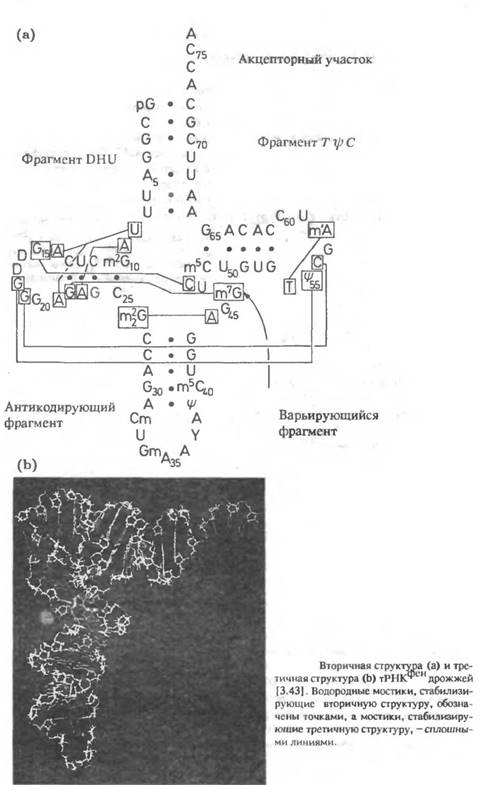
2. Исследование методом ЯМР нуклеиновых кислот и комплексов нуклеиновых кислот с протеинами
Применение метода ЯМР к исследованию нуклеиновых кислот обладает рядом особенностей по сравнению с исследованием протеинов. Так как основу нуклеотидов составляют ароматические системы, то имеет место достаточно сильная зависимость химических сдвигов от конформации молекул, что приводит к значительному усложнению спектров ЯМР. Основным фрагментом в последовательности нуклеиновых кислот является фосфатный остаток, так что ЯМР 31 С наряду с ЯМР З может быть использован в качестве достаточно чувствительного метода исследований. Ионы Mg2+ связываются с тРНК и стабилизируют структуру. На рис.3.40 показан пример спектра 31 Р тРНК при различных концентрациях Mg.
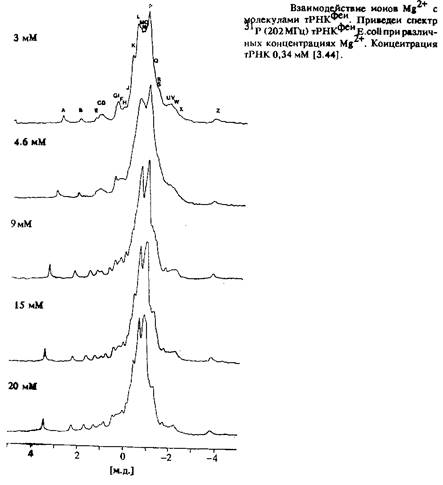
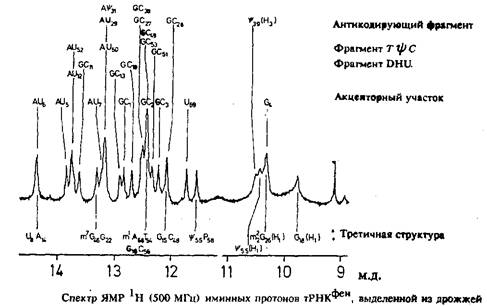
В спектре С наблюдаются хорошо разрешенные линии, положение которых изменяется в зависимости от концентрации. Даже в одномерном спектре ЯМР *Н можно достаточно четко различить отдельные линии всех протонов иминогрупп, взаимодействующих между собой, и провести их отнесение. Эти протоны имеют более важное значение для определения структуры, так как именно их взаимодействие играет основную роль при определении структуры нуклеиновых кислот.
Последовательное отнесение резонансных линий и расчет трехмерной структуры проводят теми же методами, что и при исследовании протеинов. В принципе нуклеиновые кислоты могут принимать разнообразные формы. Главная цепочка фрагмента всегда характеризуется значениями шести торсионных углов, один из которых располагается в плоскости кольца рибозы, а значит, обладает ограниченной областью значений, которая зависит от четырех эндоциклических торсионных углов. Положение основания относительно рибозы определяется одним торсионным углом в гликозидной связи. Внутри рибозы спины всех протонов связаны косвенным спин-спиновым взаимодействием. Это означает, что отнесение резонансных линий, принадлежащих рибозному остатку, может быть проведено по методу COSY. Для основания константы косвенного спин-спинового взаимодействия, как правило, невелики, так что с ростом молекулярной массы основания их далеко не всегда удается зафиксировать. Внутри основания констаиты косвенного спин-спинового взаимодействия дают неполную информацию, поэтому следует дополнительно использовать ЯЭО. В отличие от протеинов отдельные фрагменты связаны скалярным взаимодействием через фосфатные группы, и в данном случае для расшифровки структуры можно воспользоваться значением константы гетероядерного косвенного спин-спинового взаимодействия 1 ǖ 31 С.
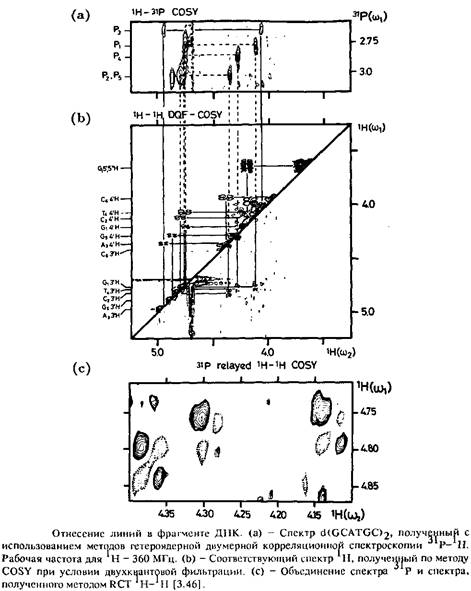
Анализ структуры резонансных линий в спектрах 31 С фосфатной группы показывает, что имеется взаимодействйе.фосфора с протоном в положении 3', с протоном в положении 5' СН2 -группы и 4' – протоном дезоксирибозы. Это означает, что проведя отнесение в спектре линии, соответствующей атому фосфора, связанному с этим протоном, можно найти линию, соответствующую протону в положении 4' следующего остатка. Правильность отнесения можно проверить, определив, какая из линий соответствует линии протона в положении 5'. Спектр З– 31 Р – J H, полученный по методу RCT, позволяет отнести линии, соответствующие протонам в положениях 3' и 4', которые принадлежат двум соседним кольцам дезоксирибозы.
Информация, получаемая из значений констант спин-спинового взаимодействия и ЯЭО, отчасти является избыточной, так что спиновая система нуклеозида может быть определена достаточно надежно. Наряду с этим имеются зависящие от пространственной структуры данные по ЯЭО для последовательности, исходя из которых можно идентифицировать соседние нуклеотиды, а также данные по сильному ЯЭО для спаренных оснований комплементарных нитей.
Расчет структуры для двойной спирали ДНК относительно прост, так как вследствие спаривания оснований и регулярности структур уже имеются достаточно надежно подтверждаемые структурные гипотезы, среди которых необходимо провести выбор наиболее вероятной и затем уточнить структуру. Подобное справедливо также и для ДНК с одной спиралью или для РНК, еслиони обладают такой структурой, как, например, тРНК.
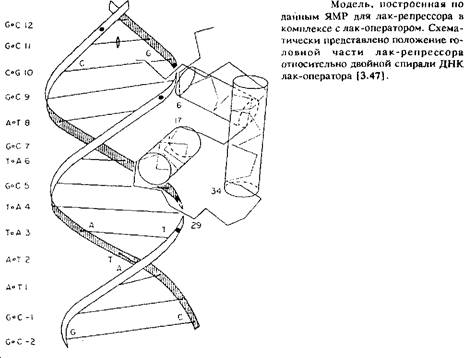
Если структура фрагмента ДНК известна и проведено отнесение резонансных линий, то можно изучить взаимодействие ДНК со специфическими протеинами. Примером этого может служить взаимодействие с протеином, который является лакрепрессором и подавляет транскрипцию лактозоспецифических ферментов, так что лактоза практически отсутствует в клетках. Структура головной части этого протеина, которая ответственна за связывание с ДНК, недавно была установлена методом ЯМР и опубликована в. По спектрам комплекса, полученным по методу NOESY, можно определить положение протеина относительно ДНК в том случае, если известно лишь несколько расстояний из измерений ЯЭО. Экспериментально по данным ЯЭО для 1 Н для трех различных ароматических колец лак-репрессора найдены структуры, соответствующие определенным нуклеотидам лак-оператора. По этим оценочным значениям расстояний получена структура комплекса, в котором лак-репрессор развернут на 180° в сравнении со структурой, полученнной из модельных представлений.
3. Состав и структура полисахаридов
Третью группу биологических макромолекул, состоящих из простых фрагментов, образуют олиго- и полисахариды. Они состоят из простых моносахаридных фрагментов, связанных между собой. Функции полисахаридов весьма разнообразны. Они играют роль резервных веществ, например крахмал, и структурных элементов, например целлюлоза. Важную функцию распознавания клеток, а также роль рецепторов выполняют поверхностные элементы: олигосахариды и малые полисахариды, связанные с липидами, свойства которых определяет их первичная структура.
Основные фрагменты полисахаридов – моносахариды – обнаруживают большее разнообразие, чем аминокислоты. Правда, моносахариды с пятью атомами углерода и с шестью атомами углерода встречаются чаще, чем другие углеводы.
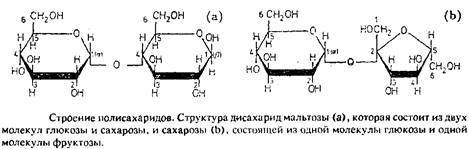
Моносахаридные фрагменты обычно связаны между собой гликозидными связями. В отличие от протеинов для них в основном характерно наличие разветвленных цепей, так как для сахаридов каждая из гидроксигрупп может образовывать точку ветвления.
4. Определение структуры полисахаридов
Химический анализ полисахаридов достаточно сложен, поэтому многие исследования сахаридов направлены на определение их химической структуры. Кольцо молекулы сахаридов может принимать различные конформации: наиболее часто встречающиеся среди них – кресло и ванна. Обе конформации характеризуются типичными значениями констант косвенного спин-спинового взаимодействия и могут достаточно просто определяться с помощью ЯМР Н. В обеих конформациях расстояния между отдельными кольцевыми протонами также будут различать совершенно определенным образом. Следовательно, и данные по ЯЭО также должны существенно отличаться. В противоположность тому, что было получено при рутинном поста-дийном определении структуры протеинов, в химии сахаридов отсутствуют простые методы последовательного определения структуры.
Для полного описания структуры необходимо знать не только картину связей моносахаридов, но и характер их связывания, т.е. необходимо дополнительно определить то, какой является связь: CL- или /3‑гликозидной. Здесь самым надежным является метод двумерного ЯМР. Если же нас интересует только картина связей моносахаридов, то отнесение сигналов в спектрах может быть существенно упрощено в случае проведения процедуры дериватизации.
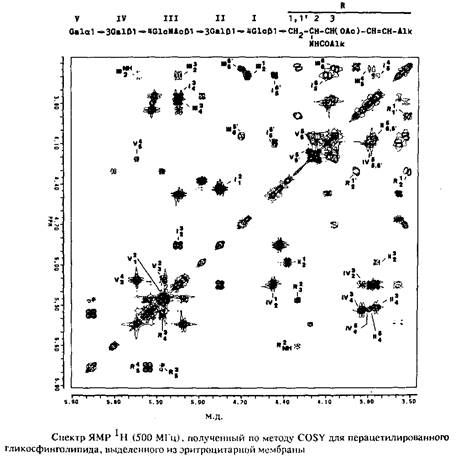
С точки зрения биологии значительный интерес представляет трехмерная структура полисахаридов, которая определяется конформацией кольцевых систем и вращением относительно простых связей в нециклических частях молекул. Принципиально структуру полисахаридов можно определить путем измерения констант связи и ЯЭО, хотя здесь могут возникнуть существенные трудности при оценке биологического соответствия определяемых структур. Связывание полисахаридов с липидами и протеинами, а также характер упаковки их в клеточные мембраны, по-видимому, оказывает сильное влияние на их пространственную структуру, так что измерения, проводимые в изолированных полисахаридах, даже при условии, что они проводятся в водной среде, необязательно должны давать реальное представление об интактной структуре. Но с другой стороны, такие измерения, по-видимому, дают наилучшее представление о структуре полисахаридов.
5. Исследование биологических мембран
Липиды образуют большой гетерогенный класс органических молекул, которые могут быть экстрагированы неполярными растворителями. Наряду с изолированными молекулами, такими, как жирорастворимые витамины и стероидные гормоны, для которых ЯМР успешно применяется для установления химической структуры, важны также фосфолипиды, которые образуют шюские, надмолекулярные структуры – двойные липидные мембраны. В соответствии с той ролью, которую играют мембраны, они были исчерпывающе исследованы с использованием всех имеющихся в распоряжении биофизических методов, включая ЯМР.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--