Курсовая работа: Исследование бактерий по почвенному профилю дерновой альфегумусовой глеевой почвы
(3,65±0,1)×109
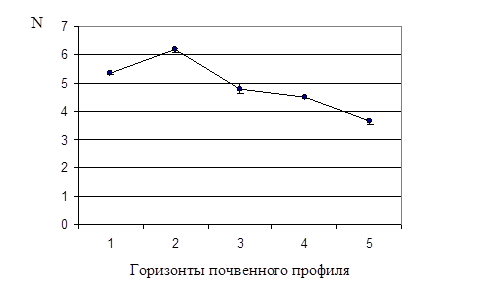
Рис. 1. Динамика общей численности бактерий в почвенном профиле дерновой альфегумусовой глеевой почвы. N – численность бактерий
(× 109 кл/г почвы).
Как видно из табл. 1 и рис. 1, максимальное значение общей численности бактерий (Nобщ. )– 6,2×109 кл/г – наблюдали во втором горизонте, по сравнению с остальными четырьмя горизонтами. Этот факт можно объяснить тем, что два верхних горизонта исследуемой почвы обладают хорошей проницаемостью для воды и поступающего с ней органического вещества. Третий горизонт почвенного профиля уже более уплотнен, поэтому органическое вещество, накапливаясь во втором горизонте, является фактором, стимулирующим развитие в нем почвенных микроорганизмов.
Данные табл. 1 показывают, что Nобщ. достоверно снижалась с увеличением глубины залегания почвенного горизонта: от второго к пятому, самому нижнему. Полученный результат согласуется с тем фактом, что содержание органического вещества, необходимого для развития почвенных микроорганизмов, также снижается с увеличением глубины залегания горизонта. При переходе от верхнего горизонта к нижележащим изменяются также окислительно-восстановительные условия, pH и др.
Порядок величин Nобщ. в каждом горизонте был одинаков – 109 кл/г. По-видимому, в верхних более рыхлых горизонтах, куда поступает достаточное количество кислорода с поверхности, создались оптимальные условия для развития аэробных микроорганизмов. В нижележащих горизонтах содержание кислорода минимально, и прежний порядок значения Nобщ. объясняется активным развитием анаэробов.
Численность сапротрофных бактерий (Nсапр. ) уменьшалась от первого горизонта к третьему, причем резкое снижение численности – более чем в 6 раз – было отмечено при переходе от первого ко второму горизонту (рис. 2А). Nсапр. (таб.2) в четвертом горизонте – 4,5×104 кл/г – была незначительно выше, по сравнению с третьим – 2,0×104 кл/г, – тогда как в пятом она опять понижалось до значения, отмеченного в третьем горизонте (рис. 2А). Как показано на рис. 2Б, численность олиготрофных бактерий (Nолиг. ) последовательно снижалась с увеличением глубины залегания почвенного горизонта от 9,5×107 в первом до 1,5×107 кл/г в пятом горизонте (таб.2).
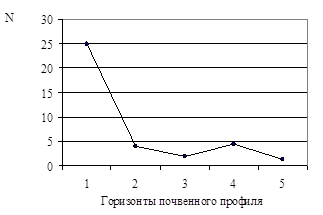
Рис. 2. Численность сапротрофов (А) и олиготрофов (Б) в почвенном профиле дерновой альфегумусовой глеевой почвы. N – численность бактерий (кл/г почвы)
Таблица 2
Численность сапротрофных и олиготрофных бактерий в почвенном профиле дерновой альфегумусовой глеевой почвы
| Горизонт | Сапротрофы | Олиготрофы |
| 1 | 2,5×105 | 9,5×107 |
| 2 | 4,0×104 | 7,5×107 |
| 3 | 2,0×104 | 3,5×107 |
| 4 | 4,5×104 | 2,0×107 |
| 5 | 1,4×104 | 1,5×107 |
В общем, Nсапр. находилась на уровне 105 кл/гв первом, и 104 кл/г в остальных горизонтах (прил. 3). Nолиг. соответствовала 107 кл/г в каждом горизонте профиля. Таким образом, Nолиг. была меньше, чемNсапр. : в первом горизонте в 100, а в остальных четырех – в 1000 раз. Выявленное значительное преобладание олиготрофов над сапротрофами свидетельствует о том, что в исследованной почве низкое содержание легкоразлагаемой органики [2], следовательно, данная почва не подвергалась антропогенному загрязнению. Бактерии функционируют в ней за счет органического вещества самой почвы. Сапротрофы, развивающиеся в условиях высокой концентрации органики [3;4], уже переработали ее большую часть. В настоящее время в исследованной почве доминируют олиготрофы, которым для существования необходимо низкое содержание органики в среде [2;4].
Интересным является факт, что для горизонтов с четвертого по пятый отмечаются признаки оглеения почвы – сизые и голубые пятна и затеки. В настоящее время известны три совокупные причины глеегенеза [5]: 1) высокая влажность; 2) наличие легкоразлагаемого органического вещества в избыточном количестве; 3) жизнедеятельность гетеротрофных микроорганизмов. Согласно нашим результатам, процесс глеегенеза идет в исследуемой почве при доминировании в микробоценозе олиготрофных бактерий, а, следовательно, в отсутствие избытка органического вещества.
Отношение Nсапр. к Nобщ. составило величины порядка 10-4 – 10-5 (табл.3), что свидетельствует о климаксном состоянии микробоценоза [3] и о доминировании бактерий, которые получают необходимые питательные вещества и энергию за счет использования трудноразлагаемых органических веществ почвы в течение длительного времени.
Таблица 3
Отношение общей численности к численности сапротрофных бактерий в почвенном профиле дерновой альфегумусовой глеевой почвы
| Горизонты (расположены от верхнего к нижнему) |
N сапр. Nобщ. |
| 1. | 0,5× 10-4 |
| 2. | 0,6×10-5 |
| 3. | 0,4×10-5 |
| 4. | 1,0×10-5 |
| 5. | 0,4×10-5 |
Примечание: Nсапр. – численность сапротрофных бактерий (кл/г); Nобщ. – общая численность бактерий (кл/г).
Заключение
1. Общая численность бактерий снижается с глубиной залегания почвенного горизонта, максимальное значение – 6,2×109 кл/г – наблюдается во втором горизонте за счет того, что два верхних горизонта исследуемой почвы обладают хорошей проницаемостью для воды и поступающего с ней органического вещества.
2. Количество сапротрофных бактерий уменьшалось от первого горизонта к третьему – от 2,5×105 до 1,4×104 кл/г, – причем резкое снижение численности – более чем в 6 раз (от 2,5×105 до 4,0×104 кл/г) – было отмечено при переходе от первого ко второму горизонту.
3. Численность олиготрофных бактерий последовательно снижалась с увеличением глубины залегания почвенного горизонта от 9,5×107 в первом до 1,5×107 кл/г в пятом горизонтах.
4. Отношение численности олиготрофных бактерий к сапротрофным для первого горизонта составляет величину порядка 102 , а в остальных четырех – 103 , что свидетельствует о значительном преобладании олиготрофов над сапротрофами.
5. Отношение численности сапротрофных бактерий к общей численности бактерий составляет величины порядка 10-4 – 10-5 , что свидетельствует о климаксном состоянии микробоценоза и о доминировании бактерий, жизнедеятельность которых связана с использованием трудноразлагаемых органических веществ почвы.
6. Согласно нашим результатам, процесс глеегенеза идет в исследуемой почве при доминировании в микробоценозе олиготрофных бактерий, а, следовательно, в отсутствие избытка органического вещества.
Список использованных источников и литературы
1. Бабьева И.П. Биология почв / И.П. Бабьева, Г.М. Зенова. – М.: МГУ, 1989. – 336 с.
2. Емцев В.Т. Микробиология / В.Т. Емцев, Е.Н. Мишустин. – М.: Дрофа, 2005. – 445 с.
3. Добровольская Т.Г. О показателях структуры микробных сообществ / Т.Г. Добровольская, И.Ю. Чернов, Д.Г. Звягинцев // Микробиология. – 1997. – Т.66. – №3. – С. 408–414.
4. Заварзин Г.А. Введение в природоведческую микробиологию / Г.А. Заварзин, Н.Т. Колотилова. – М.: Книжный дом «Университет», 2001. – 256 с.
5. Зайдельман, Ф.Р. Процесс глееобразования и его роль в формировании почв / Ф.Р. Зайдельман. – М.: МГУ, 1998. – 316 с.
6. Кузнецов С.И. Методы изучения водных микроорганизмов / С.И. Кузнецов, Г.А. Дубинина. – М.: Наука, 1989. – 288 с.
7. Практикум по микробиологии / Под ред. Н.С. Егорова. – М: МГУ, 1976. – 307 с.
8. Соединения железа, алюминия, кремния и марганца в почвах лесных экосистем таежной зоны / М. Мурашкина, Г. Копцик, и др. // Почвоведение. – 2004. – № 1. – С. 40– 49.
9. Умаров М.М. Экология и почвы / М.М. Умаров. Избранные лекции 1–7 школ, М.: Пущино, 1998, 15–21 с.
10. Экология микроорганизмов: учебник / А. И. Нетрусов, Е.А. Бонч - Осмоловская, и др. – М.: Издательский центр «Академия», 2004. – 272с.
11. Hottes A.K. Transcriptional profiling of Caulobacter crescentus during growth on complex and minimal media / A.K. Hottes, M. McEwen, D. Yang et al. // J. Bacteriol. – 2004. – V. 186. – №5. – P. 1448–1461.