Курсовая работа: Цитология эукариотических водорослей
Протопласт как совокупность живых компонентов клетки лишь у сравнительно немногих более примитивных водорослей ограничен снаружи только цитоплазматической мембраной. Такие клетки называют «голыми». Голыми являются, как правило, репродуктивные клетки водорослей – зооспоры и гаметы.
Клеточная оболочка
У большинства водорослей протопласты полностью окружены клеточными стенками. Как и у высших растений, клеточная оболочка водорослей всегда образуется кнаружи от плазмалеммы, защищает протопласт и служит как бы наружным скелетом клетки. Оболочка состоит из двух компонентов: аморфного слизистого матрикса и погруженного в него фибриллярного опорного компонента. В состав матрикса входят гемицеллюлозы и пектиновые вещества, образуемые аппаратом Гольджи внутри клетки и выделяемые через плазмалемму путем экзоцитоза. Что касается химического состава фибриллярного компонента клеточных стенок, го в отличие от высших растений, где фибриллы представлены целлюлозой, у водорослей наблюдается большее разнообразие полисахаридов, образующих скелетные фибриллы. «Создается впечатление, что природа экспериментировала с разнообразными типами полисахаридов до тех пор, пока отбор не показал, наконец, что именно цепи целлюлозы могут служить наилучшим материалом для укрепления клеточных оболочек в борьбе растений за переход к наземному образу жизни». К настоящему времени у водорослей на молекулярном и ультраструктурном уровне детально описаны три главных типа скелетных полисахаридов: целлюлоза, манпап и ксилан.
Целлюлоза обнаружена у многих зеленых водорослей, у желто-зеленых, динофитовых. в небольших количествах – у красных и бурых.
Ряд зеленых водорослей с сифоновой организацией таллома не содержит в стенках целлюлозы. Главные составные части клеточных стенок у них – ксилан или маннан. Так, ксилан обнаружен в клеточных стенках бриопсиса, каулерпы, галимеды, удотеи, а маннан – у ацетабулярии и кодиума. Это различие в химическом составе оболочек пытались использовать для установления родственных связей в пределах этой группы зеленых водорослей. Предлагалось делить их на «ксилановую» и «манпаповую» группы. Однако позднее ксилан и маннан были обнаружены в стенках одних и тех же водорослей на разных стадиях их жизненного цикла: спорофиты дертэезии, бриопсиса, псевдобриопсиса содержали маннан, а гаме тофиты – ксилан и целлюлозу. В ряде случаев состав микро-фибрилл точно не установлен.
Фибриллы – скелетные волокна – ориентированы в матриксе по-разному. Они могут быть рассеяны но нему беспорядочно или же они располагаются параллельно друг другу. И то и другое расположение микрофибрилл может встречаться в пределах одной систематической группы, в разных частях одного и того же таллома и даже в разных слоях клеточной стенки одной и той же клетки.
Например, среди зеленых водорослей беспорядочное расположение фибрилл наблюдается у родов, клеточные стенки которых характеризуются обильным аморфным матриксом. У сифонокладальных стенка содержит лишь небольшие количества аморфного материала. Она состоит из многочисленных ламелл, в которых фибриллы ориентированы более или менее параллельно друг другу. В соседних ламеллах фибриллы располагаются почти под прямым углом.
Первый слой новой стенки, образующийся на остановившихся зооспорах этих водорослей, состоит из беспорядочно расположенных микрофибрилл.
Помимо скелетных полисахаридов, матрикс клеточных стенок может содержать инкрустирующие вещества – кремнезем, спорополленин, карбонат кальция. Есть данные о присутствии в клеточных стенках протеинов. В 1972 г. у ряда видов хлореллы, сценедесмуса, прототеки в их оболочках впервые был обнаружен спорополленин – чрезвычайно стойкое по отношению к ферментам вещество, характерное для оболочек спор и пыльцевых зерен высших растений. За последние годы число водорослей, в оболочках которых найден спорополленин, заметно возросло. Он обнаружен в клеточных стенках Phycopeltiseplphiton, ряда видов требуксии, в стенках цист гемато-кокка, в стенках зигот спирогиры. У некоторых водорослей имеется хитин в виде тонкого наружного слоя. Оболочка может быть цельной или состоять из двух и более частей, может быть пронизана порами, нести различные выросты. Примеры будут приведены при описании соответствующих родов.
Формирование новых оболочек исследовалось с применением различных методов электронной микроскопии. Объектами служили голые, одетые только цитоплазматической мембраной естественные стадии развития разных водорослей: цисты ацетабулярии, только что оплодотворенные яйцеклетки фукуса, синзооспоры вошерии, зооспоры педиаструма. кладофоры, хетоморфы; автоспоры хлореллы, ооцистиса, сценедесмуса. В других исследованиях по регенерации стенки использовались искусственно полученные протопласты. Результаты этих исследований несколько противоречивы; по-видимому, сборка клеточных стенок происходит неодинаково у разных водорослей, и в этот процесс вовлечены разные органеллы.
В ряде случаев констатируется участие в синтезе клеточных стенок аппарата Гольджи после остановки зооспор возникает оболочка. При этом на плазмалемме этих голых клеток видны гранулы с той же ориентацией, что и микрофибриллы в зрелых оболочках клеток этих водорослей. Был сделан вывод, что эти гранулы плазмалеммы участвуют в биосинтезе целлюлозных микрофибрилл. Сходные данные были получены для автоспор Oocystisapiculata– водоросли, также имеющей упорядоченную текстуру микрофибрилл в оболочке зрелых клеток. Исследование реплик плазмалеммы голых автоспор этой водоросли показало, что снаружи плазмалемма несет спаренные ряды мелких гранул, соединяющиеся в широкие полосы. На конце рядов может находиться более крупная гранула. Между полосами гранул располагаются отдельные гранулы разного диаметра. Ориентация полос гранул точно соответствует ориентации микрофибрилл в оболочке.
С внутренней стороны плазмалемма несет отдельные гранулы. Было высказано предположение, что полосы гранул являются местом синтеза целлюлозных микрофибрилл, которые вставляются в глубокую выемку между парами рядов гранул. С другой стороны, из анализа электронных микрофотографий кортикального слоя цитоплазмы и клеточной стенки Oocystissolitariaсделан вывод, что упорядоченная организация микрофибрилл обусловлена микротрубочками, сосредоточенными в примембранном слое цитоплазмы. В пользу этой гипотезы свидетельствуют опыты с ингибиторами, действующими на микротрубочки, например с колхицином. Обработка клеток колхицином приводит к деполимеризации микротрубочек и нарушает ориентацию микрофибрилл. У водорослей, помимо цитоплазматической мембраны и клеточных стенок, наблюдается большое разнообразие клеточных покровов. Эти покровы можно распределить на две группы: формирующиеся под цитоплазматической мембраной и находящиеся кнаружи от нее. К первым относятся: перипласт криптофитовых, пелликула эвгленофитовых и тека динофлагеллат; ко вторым – различного рода чешуйки, образующие покров снаружи от цитоплазматической мембраны, особенно характерные для золотистых и гаптофициевых водорослей, но встречающиеся и у зеленых. Особые покровы – панцири диатомовых водорослей, неполные оболочки–домики золотистых, эвгленофитовых водорослей.
Цитоплазма
Цитоплазма – часть клетки, остающаяся после удаления различимых в световом микроскопе органелл. Основная плазма – мелкогранулированный матрикс – содержит ультраструктурные органеллы цитоплазмы: плазмалемму, эндоплазматическую сеть, аппарат Гольджи, микротрубочки и пр.
Мембраны. Электронный микроскоп показывает, что цитоплазма пронизана многочисленными мембранами, имеющими на срезе вид двойных темных линий, заключающих светлое промежуточное пространство. Снаружи клетка ограничена такой типичной элементарной, мембраной – плазмалеммой, которая в живой клетке обладает свойством полупроницаемости и выполняет барьерную функцию. Все клеточные органеллы ограничены такой элементарной мембраной. Ядра, пластиды, митохондрии отделены от основной плазмы оболочками, состоящими из двух мембран.
Внутри клетки цитоплазма отграничена от вакуолей вакуолярной мембраной – тонопластом, имеющим ту же природу, что и плазмалемма. Наряду с перечисленными стабильными мембранами, отграничивающими клетку от наружной среды и органеллы от основной плазмы, существует еще чрезвычайно изменчивая внутренняя система мембран – эндоплазматическая сеть. Эта система образована ограниченными элементарной мембраной плоскими полыми цистернами и трубочками, связанными в сеть. Наружная поверхность эндоплазматической сети часто покрыта рибосомами. Соответственно различают шероховатую и гладкую эндоплазматическую сеть, которая может встречаться в одной и той же клетке. Эндоплазматическая сеть играет роль в передвижении и распределении веществ внутри клетки. Рибосомы, связанные с мембранами шероховатой эндоплазматической сети, осуществляют синтез белка.
Аппарат Гольджи. Аппарат Гольджи состоит из диктиосом, одной или многих. Каждая диктосома представляет собой стопку плоских блюдцевидных цистерн, ограниченных элементарной мембраной. Число цистерн в стопке, т.е. приходящихся на одну диктиосому, варьирует от 3–7 до 30. От краев цистерн, большей частью вздутых, отшнуровываются мелкие пузырьки – везикулы Гольджи. Вся органелла носит полярный характер; в ней имеются проксимальный, или формирующий, полюс и дистальный, или секретирующий, полюс, где цистерны образуют обильные пузырьки и фрагментируются. Таким образом, цистерны постоянно перемещаются от формирующего полюса к секретирующему. Образование новых цистерн Гольджи, по-видимому, происходит за счет слияния пузырьков, которые отпочковываются от эндоплазматической сети или наружной мембраны ядерной оболочки. Если основной функцией эндоплазматической сети является полимеризация аминокислот в специфические полипептиды, то в аппарате Гольджи из простых Сахаров синтезируются олигомерные углеводы. По мере созревания цистерн диктиосомы водянистая жидкость эндоплазматической сети или перинуклеарного пространства, как правило, обогащается углеводами, что можно выявить гистохимически. Функции аппарата Гольджи многообразны. Он участвует в образовании матрикса клеточной оболочки. В результате слияния пузырьков Гольджи возникает клеточная пластинка, принимающая участие в цитокинезе многих зеленых водорослей. Срединная пластинка представляет собой полужидкий слой из гемицеллюлозы и пектиновых веществ, составляющий матрикс клеточной оболочки, в котором позднее появляются фибриллы целлюлозы. Синтез фибрилл, как правило, осуществляется плазмалеммой. Мембраны же пузырьков Гольджи преобразуются в плазмалемму. Аппарат Гольджи поставляет также материал матрикса для растяжения клеточной стенки. Пузырь, где откладывается кремнезем и в котором формируется новая створка диатомовых водорослей, также возникает за счет слияния пузырьков Гольджи. То же относится и к мембране – силиколемме, окружающей пузырь, в котором закладывается кремнеземная оболочка эндоплазматических статоспор золотистых водорослей. В цистернах аппарата Гольджи наблюдаются не только растворенные вещества, но и оформленные чешуйки, составляющие клеточный покров многих монадных золотистых и некоторых зеленых водорослей. Чешуйки выделяются на поверхность клетки путем экзоцитоза - при слиянии мембраны цистерны, содержащей готовую чешуйку, с плазмалеммой. В этой связи заслуживают внимания данные Р. Броуна с сотрудниками, относящиеся к гаптофициевой водоросли – PleurochrysisscherffelLНа поперечном разрезе через зрелую вегетативную клетку этой водоросли можно видеть, что клеточный покров образован многими слоями рыхло расположенных чешуек, погруженных в материал, по-видимому, пектиновой природы. На ташентальном срезе клетки чешуйки представляются в плане: их остов образован целлюлозными микрофибриллами, из которых одни располагаются концентрическими кругами, другие расходятся радиально из центра чешуек. Эти чешуйки формируются в цистернах аппарата Гольджи. Процесс начинается в проксимальных цистернах: центральная расширенная часть таких цистерн является местом полимеризации глюкозных мономеров в кристаллическую сеть целлюлозных цепей. В дистальных цистернах можно видеть уже зрелые чешуйки; их остов образован сетью целлюлозных микрофибрилл, одна часть которых расположена концентрическими кругами, а другая расходится из центра по радиусам. Мембраны самых наружных цистерн с уже готовыми чешуйками, по-видимому, сливаются с цитоплазматической мембраной, принимая, таким образом, участие в выделении готовых элементов клеточного покрова па поверхность клетки, где они располагаются наподобие черепиц. Подсчитано, что у Pleurochrysisscherffeliаппарат Гольджи каждые две минуты продуцирует одну чешуйку. Таким образом, скорость продукции чешуек достаточно велика, что, возможно, связано с большой потерей рыхло расположенных чешуек.
Было доказано, что основной строительный полисахарид чешуек плеврохризиса – целлюлоза. Этим было продемонстрировано, что аппарат Гольджи участвует не только в синтезе аморфного матрикса клеточной стенки, но и в синтезе целлюлозы, принимая на себя морфогенетические функции, обычно выполняемые плазмалеммой. До этого исследования клеточного покрова P. scherffeliобщепризнанным считалось, что опорные фибриллы целлюлозы образуются всегда вне протопласта благодаря направленной наружу активности плазмалеммы. Возникновение чешуек, кокколитов в цистернах аппарата Гольджи к настоящему времени описано у многих представителей.
В ряде случаев, например у золотистой водоросли охромонас, было показано, что в сборку и выведение на поверхность клетки мастигонем вовлечены пузырьки Гольджи. Сами мастигонемы, будучи белковой природы, образуются не в пузырьках Гольджи, а в перинуклеарном пространстве, т.е. в системе синтезирующей белок эндоплазматической сети.
Одна из функций аппарата Гольджи – регуляция содержания воды в клетке. У рафидофитовой водоросли Vacuolariavirescensбыло продемонстрировано возникновение пульсирующих вакуолей из аппарата Гольджи. Сократительная вакуоль этой водоросли представляет собой разбухшую цистерну Гольджи. При «сокращении» она сливается с плазмалеммой, и ее жидкое содержимое удаляется из клетки путем экзоцитоза. Вакуоль, по существу, и не является сократительной, так как она полностью исчезает и заменяется новой цистерной. Это процесс, обратный процессу активного поглощения воды путем пиноцитоза. Было показано, что каждые 75 с из клетки выталкивается новая вакуоль и за 30 мин выделяется количество воды, соответствующее полному объему клетки. Аппарат Гольджи связан с секреторной деятельностью клетки, при его участии образуются, по-видимому, трихоцисты, эджектосомы, возможно, выделяется клеющая субстанция, способствующая прикреплению спор к субстрату, и т.д.
Микротрубочки. Помимо эндоплазматической сети, в цитоплазме клеток водорослей обнаруживаются трубчатые элементы – цитоплазматические микротрубочки. Их следует отличать от микротрубочек, составляющих аксонему ресничек и жгутиков. В отличие от последних образование цитоплазматических микротрубочек подавляется действием колхицина, они исчезают из цитоплазмы при низкой температуре и высоком гидростатическом давлении. Цитоплазматические микротрубочки–крайне динамические клеточные структуры: они то появляются, то исчезают, уменьшаются или увеличиваются в числе, перемещаются из одной части клетки в другую. Стенки микротрубочек образованы глобулярными макромолекулами. Хотя цитоплазматические микротрубочки пока не удается выделить из цитоплазмы и анализировать, на них обычно распространяются данные, полученные в отношении микротрубочек жгутиков, поддающихся изолированию и биохимическому анализу. Принимается, что цитоплазматические микротрубочки, как и микротрубочки жгутиков, образованы белками тубулинами, по аминокислотному составу напоминающими актин мышц. Цитоплазматические трубочки различаются по положению в клетке и назначению: одни участвуют в образовании ядерного веретена; другие связаны с цитокинезом – участвуют в образовании фрагмопласта и фикопласта; третьи, собственно цитоплазматические микротрубочки, представляют собой скелетную систему клетки. Последняя особенно свойственна голым клеткам, форма которых определяется микротрубочками, образующими периферический микроскелет. Например, асимметричная форма клетки голой золотистой водоросли охромонаса определяется присутствием цитоплазматических микротрубочек: при высоком гидростатическом давлении или обработке колхицином клетки охромонаса принимают сферическую форму. Своеобразная тетраэдрическая форма зооспор золотистой водоросли гидруруса также определяется системой микротрубочек. Микротрубочки ответственны не только за поддержание, но и за изменение формы голых клеток, например зооспор водяной сеточки, педиаструма во время формирования их дочерних ценобиев, а также в процессе инверсии при образовании дочерних сфероидов вольвокальных. Микротрубочки, локализованные на переднем конце сперматозоидов эдогониума, возможно, наряду со жгутиковыми корнями и фиброзным кольцом ответственны за те изменения формы кончиков сперматозоидов, которые наблюдаются перед самым процессом плазмогамии. В ряде случаев микротрубочки наблюдались по соседству с хлоропластами, текальными цистернами у динофитовых водорослей, пелликулой эвгленофитовых и пр.
Жгутики
Строение жгутиков. Монадные клетки снабжены жгутиками. Жгутики всех водорослей имеют единый план строения.
Жгутик можно подразделить на наружный и внутренний – эстра- и интрацеллюлярный отрезки. Наружный отрезок, или свободная часть жгута, в свою очередь, состоит из трех частей: кончика, главного стержня и переходной зоны. Внутренний - интрацеллюлярный – отрезок охватывает основание жгута и корни жгута.
Свободная часть жгута всегда одета жгутиковой мембраной, которая не что иное, как приподнятая цитоплазматическая мембрана. Так, у бурой водоросли диктиоты единственный жгутик несколько раз свернут по спирали вокруг молодого сперматозоида. На срезе через такой сперматозоид под мембраной, одевающей клетку, можно видеть микротрубочки жгута, характерным образом расположенные, без какой-либо собственной мембраны. После освобождения сперматозоида первоначально закрученный по спирали жгутик разматывается, приподнимая при этом цитоплазматическую мембрану клетки, которая на этой стадии должна быть в очень лабильном состоянии. Наоборот, при втягивании жгутиков, например при остановке синзооспор вошерии, их мембрана становится частью цитоплазматической мембраны. Кнаружи от жгутиковой мембраны находится чехол – слой материала, покрывающего жгут на большем протяжении его длины.
Под мембраной жгута находится матрикс, образованный белками. Весь жгутиковый аппарат пронизан пучком белковых фибрилл или микротрубочек – аксонемой, поперечный разрез которой выглядит неодинаково на разных уровнях жгута: в его кончике, главном стержне, переходной зоне и базальном теле.
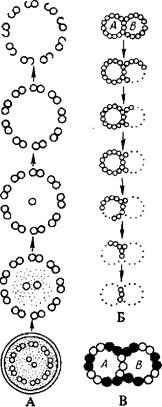
На поперечном срезе главного стержня видно типичное для всех водорослей расположение микротрубочек или фибрилл: дольцо из девяти пар микротрубочек окружает две одинарные центральные микротрубочки – структура +2.Каждый дублет образован «А» и «В» микротрубочками, «А» трубочка каждого дублета снабжена двумя короткими выступами–боковыми «руками», обращенными к следующему дублету, и радиальным тяжем, соединяющим периферический дублет с центральными микротрубочками, которые могут быть заключены в общий чехол. Кроме того, имеются интердублетные соединения – тяжи, соединяющие дублеты, и ряд дополнительных образований.
А последовательность растворения микротрубочек жгутика: удаление мембраны жгутика оставляет нетронутой аксонему. содержащую девять наружных дублетов и две центральные трубочки. При обработке аксонемы возрастающими концентрациями саркозила удаляется одна из центральных трубочек, затем растворяется оставшаяся центральная трубочка, часто вместе с наружной частью трубочки В. Б- интактные наружные дублеты растворяются в такой последовательности: наружная часть трубочки В, оставшаяся часть трубочки В, наружная часть трубочки А, А–остаток трубочки А, за исключением грех первичных нитей, составляющих перегородку между трубочками А и В. В схема, иллюстрирующая гипотетическое расположение тубулина 1 и тубулина 2 в наружном дублете.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--