Реферат: Генная инженерия 4
Источник экзогенной ДНК не имеет значения. ДНК может быть получена, например, из клеток человека, но можно сшивать и искусственно синтезированные гены. Кроме бактериальных плазмид в качестве векторов (носителей) ДНК используют фаги λ (объект исследования Альберта). Часть генома этого фага не обязательна для его размножения в бактерии. Вместо него можно ввести чужеродную ДНК, которая будет размножаться вместе с фаговой после инфицирования бактерий.
Добиться репликации и амплификации в составе плазмидной (или фаговой) ДНК после трансформации бактериальной клетки ещё не значит решить все её проблемы. Прежде всего возникают два вопроса:
1. Как распознавать клоны, содержащие гибридную ДНК, среди потомства трансформированных клеток или живых бактериофагов ?
2. как идентифицировать необходимые фрагменты ДНК среди многих клонированных неизвестных фрагментов?
Например можно отбирать бактериальные клетки, если они несут плазмиду с фактором устойчивости к антибиотику, выращивая их на среде, на среде, содержащей антибиотик. Нетрансформированные клетки без плазмид(и, следовательно, без гена устойчивости к антибиотику) просто не будут расти на такой среде. В последнее время разработано много специальных методов вакцинации, которые позволяют отбирать только рекомбинантные клетки.
Для генной инженерии белков недостаточно отобрать и размножить определённые фрагменты ДНК, необходимо ещё индуцировать их экспрессию в клетке. Для этого необходимо «подключить» рекомбинантную молекулу ДНК , последующую трансляцию матричной РНК и процессинг как на транскрипционном , так и на трансляционных уровнях.
3.3. Идентификация и анализ генов
Ещё одна область применения рестриктаз – идентификация и определение числа генов. Эти задачи решаются с помощью метода разработанного Саузерном.
Тотальную ДНК из клеток человека гидролизуют эндонуклеазой примерно на 500000 фрагментов длиной от 102 до 105 нуклеотидных пар. Затем фрагменты разделяют по молекулярной массе с помощью гель- электрофореза в ага розе, после чего ДНК денатурирует с щелочью прямо в геле, чтобы получить одноцепочные фрагменты. Их переносят на нитроцеллюлозный фильтр и фиксируют высушиванием при 800 С. В результате получается отпечаток(реплика) картины разделения фрагментов ДНК по их размеру. Эти фрагменты можно идентифицировать методом гибридизации с радиоактивными ДНК-зондами, специфичными для определённых генов или хромосом. Любой фрагмент, содержащий всю последовательность зондируемого гена или его часть, будет выглядеть на радиоавтографе в виде тёмной полосы.
Зонды и генные библиотеки. Главное условие такого анализа - наличие подходящего геноспецифического радиоактивного ДНК-зонда, который можно использовать для гибридизации.
3 .4. Сортировка хромосом
Следующий метод – это метод сортировки хромосом при помощи цитофлюрометрии. Этот метод может быть использован в двух разных целях:
1) Для идентификации и количественного анализа большого числа хромосом в течение очень короткого времени.
2) Для препаративного разделения хромосом. Этот метод имеет два преимущества перед стандартными методами анализа хромосом:
- во-первых, он полностью автоматический, благодаря чему исключается элемент субъективности
- во-вторых, он намного быстрее (рис. 3)
Однако важнее, что этот метод позволяет препаративно разделять хромосомы, и при наличии специфических зондов исследовать структуру и функцию отдельных генов становится относительно просто. В этом случае ген можно локализовать в хромосоме с помощью гибридизации in situ, размножить его ДНК путём клонирования и секвенирования.
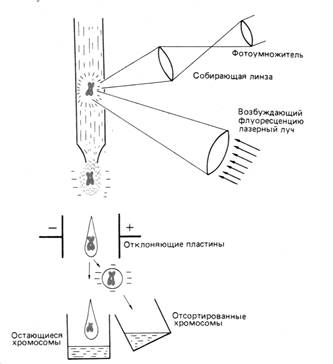 |
Рис. 3 Принцип сортировки хромосом с помощью лазера. Хромосомы окрашены флуоресцирующим красителем. Флуоресценция возбуждается лазерным лучом и измеряется для каждой хромосомы отдельно. Данные измерений используют для сортировки хромосом.
3.5. Секвенирование ДНК
Последовательность нуклеотидов и генетический код.
Методы определённой последовательности аминокислот в полипептидной цепи были известны ещё в 50-х годах. Теоретически это относительно лёгкая проблема, поскольку все 20 аминокислот, встречающихся в природных белках, имеют разные свойства. С другой стороны, нуклеотидная последовательность ДНК относительно однородна по составу однородных звеньев, так как содержит только четыре типа азотистых оснований – гуанин, цитозин, аденин и тимин. Когда в 60-е годы был расшифрован генетический код, появилась возможность востанавливать (дедуцировать) нуклеотидную последовательность соответствующего белка. Однако генетический код является вырожденным, то есть одной и той же аминокислоте соответствуют несколько нуклеотидных триплетов. Следовательно сведения о нуклеотидной последовательности аминокислот в белке, не однозначны. Кроме того последовательности аминокислот не содержат никакой информации о последовательности некодирующих участков ДНК. Принцип состоит в следующем: длинную молекулу ДНК фрагментируют при помощи агентов, расщепляющихся в специфических сайтах. Затем определяют последовательность нуклеотидов в каждом из этих фрагментов. Очерёдность фрагментов в целой молекуле восстанавливают, используя перекрывающие концы: идентичные цепи разрезают повторно другой рестриктазой, а затем последовательности, перекрывающихся образующихся при обработке двумя рестриктазами разной специфичности, сравнивают. Так может быть реконструирована полная последовательность. В пределах отдельных фрагментов порядок нуклеотидов определяют с помощью специальных методов. Раньше секвенирование ДНК было весьма трудным делом, теперь же оно осуществляется очень легко и быстро.
3.6. Динамичность генома
Методы новой гентики расширили наши знания о структуре генетического материала. В 1963 году Тэйлор описал “индуцированные фагом мутации E. Coli”, вкоре после этого, Старлингер и Седлер описали IS-элементов у бактерий. Эти элементы получили название мобильных, теперь же они определяются как специфические последовательности ДНК, которые могут неоднократно внедряться в разные сайты генома. Перенос генов от одной бактерии к другой с помощью фага (трансдукция) известен давно, а теперь используется и в генетической инженерии эукариот (включая клетки млекопитающих). Возможно, такие процессы могут происходить и в природе. Более того, последовательности ДНК, гомологичные глобиновому гену человека, были обнаружены у бобовых растений. Функция такого гена у растений может состоять в том, чтобы “обеспечить кислородом клубеньковые бактерии в ткани”. Наличие этого гена может быть объяснено переносом его от насекомых или млекопитающих.
4. Области практического применения генной инженерии
4.1. Создание трансгенных растений
Еще 10 лет тому назад биотехнология растений заметно отставала в своем развитии, но за последние 3 года наблюдается быстрый выброс на рынок трансгенных растений с новыми полезными признаками. Трансгенные растения в США в 1996 году занимали площадь 3 млн. акров, в 1997 году площадь увеличилась до 15 млн. акров, в 1998 году – до 60 млн. акров, а в прошлом году до 80 млн. акров. Поскольку основные трансгенные формы кукурузы, сои, хлопчатника с устойчивостью к гербицидам и насекомым хорошо себя зарекомендовали, есть все основания ожидать, что площадь под генноиженерные растения в будущем (2001 году) увеличатся в 4-5 раз.
В апреле 1998 года доля в процентах трансгенных форм растений в сельском хозяйстве составило:
- кукуруза – 6
- соя – 12
- хлопчатник – 15
- томаты – <1
Так как число жителей за последнее столетие увеличилось с 1.5 до 5.5 млрд. человек, а к 2020 году предполагается вырост до 8 млрд., таким образом возникает огромная проблема, стоящая перед человечеством. Эта проблема заключается в огромном увеличение производства продуктов питания, несмотря на то, что за последние 40 лет производство увеличилось в 2.5 раза, все равно этого не достаточно. И в мире в связи с этим наблюдается социальный застой, который становится все более настоятельным. Другая проблема возникла с медицинским лечением. Несмотря на огромные достижение современной медицины, производимые сегодня лекарственные препараты столь дороги, что ¾ населения земли сейчас полностью полагаются на традиционные донаучные методы лечения, прежде всего на неочищенные препараты растительного происхождения.