Реферат: Хромосоми як матеріальна основа спадковості. Зміни хромосом та механізми їх реорганізації
2.1 Мітоз
Мітоз, або непряме ділення, - основний спосіб розмноження еукаріотичних клітин, що обумовлює, зокрема, можливість збільшення їх біомаси, зростання і регенерацію. Мітоз складається з чотирьох фаз.
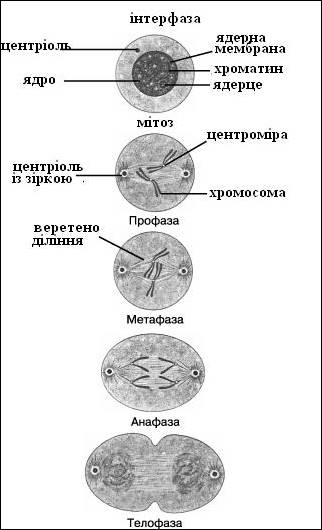
Перша – профаза – характеризується початком циклу компактизації хромосом, який продовжується протягом всієї цієї фази. Внаслідок цього хромосоми стають видимими під мікроскопом, причому вже в середній профазі мітоза вони представляються подвійними структурами – сестринськими хроматидами, закрученими одна довкола іншої. До кінця профази зникають ядерце і ядерна мембрана.
Друга –метафаза. Процес компактизації хромосом продовжується і веде до ще більшого укорочення їх довжини. Хромосоми вишиковуються по екватору клітки. Хроматиди сполучені між собою між собою в центромері, званою також первинною перетяжкою. З'являються нитки мітотичного веретена, які приєднуються до ценромерів. Кожна ценромера випробовує напругу, оскільки нитки веретена тягнуть її до протилежних полюсів.
Полюси клітки формуються спеціальними органелами – центросомами.
Третя – анафаза – починається з розриву ценромери, внаслідок чого сестринські хроматиди розходяться до різних полюсів клітки. З цієї миті кожна пара сестринських хроматид отримує назву дочірніх хромосом.
Четверта – телофаза. Хромосоми досягають полюсів клітки, з'являються ядерна мембрана, ядерце. Відбуваються декомпактізация хромосом і відновлення структури інтерфазного ядра. Закінчується мітоз діленням цитоплазми і в типових випадках – відновленням вихідної біомаси дочірніх клітин.
Біологічна роль мітоза полягає в забезпеченні ідентичною генетичною інформацією двох дочірніх кліток. Це досяжно лише завдяки циклу компактізациі – декомпактізациі, який і дозволяє розподілити спадкові молекули в мінімальному об'ємі мітотичних хромосом. Інакше, враховуючи розміри клітки (десятки або сотні кубічних мікрометрів) і довжину декомпактізованной хромосоми (сантиметри), кожне клітинне ділення супроводилося б хаотичним переплетенням хромосомного матеріалу.
У еволюції еукаріотичних клітин, мабуть, ця обставина і послужила причиною становлення настільки складного генетичного процесу, як мітоз.
2.2 Мейоз
Терміном «мейоз» позначають два наступних один за одним ділення, в результаті яких з диплоїдних кліток утворюються гаплоїдниє статеві клітини, – гамети. Якби запліднення відбувалося диплоїдними гаметамі, то плодоїдність нащадків в кожному наступному поколінні повинна була б зростати в геометричній прогресії. В той же час завдяки мейозу зрілі гамети завжди гаплоїдни, що дозволяє зберігати діплоїдность соматичних кліток виду. Можливість існування подібного до мейозу ділення при дозріванні гамет тварин і рослин була передбачена А. Вейсманом ще в 1887 р. Мейотичне ділення не еквівалентне мітозу. Обом мейотичним діленням передує лише одна фаза синтезу ДНК.

Тривалість її, як і профази I ділення мейозу, у багато разів перевершує відповідні показники мітотичного циклу будь-яких соматичних клітин даного виду. Головні події мейозу розгортаються в профазі I ділення. Вона складається з п'яти стадій.
У першій стадії – лептотені, слідуючою безпосередньо за закінченням предмейотичного синтезу ДНК, виявляються тонкі довгі хромосоми. Вони відрізняються в профазі мітоза двома особливостями: по-перше, в них не виявляється подвійність, тобто не видно сестринських хроматид, по-друге, лептотенні хромосоми мають виражену хромомерну будову. Хромомери – вузлики. Ділянки щільної компактізациі ДНК, розміри і розташування яких строго видоспецифічний. Хромомери зустрічаються як в мейотичних, так і в мітотичних хромосомах, проте в останніх без специфічної обробки вони не помітні.
Третя стадія профази I ділення – пахитена – у більшості видів найтриваліша. Під світловим мікроскопом видно кон’югіровавші хромосоми з більш менш чітко вираженою хромомерною будовою. Приблизно в середині пахитени між хроматидами гомологічних хромосом з'являється подовжня щілина, яка чітко показує, що бівалент – це, по суті, четверна хромосомна структура. У пахитене відбувається важлива генетична подія – кросинговер, або перехрещення хроматид гомологічних хромосом. В результаті цього в кожному гомологу змішуються батьківський і материнський спадковий матеріал.
Результати кросинговера стають помітними лише в четвертій і п'ятій стадіях профази I ділення – діплотені і діакенезі. Діплотена починається з моменту розбіжності гомологічних хромосом. В цей час в точках кросинговера видно перехрещені хроматиди. Область перехрещення хроматид називають хіазмою. Число хіазм в цілому відповідає кількості актів кросинговера в біваленті і пропорційно довжині гомологічних хромосом, його складових. Для діплотени і діакенезу характерне прогресуюче укорочення хромосом в результаті компактізаціі; тому хіазми поступово терміналізуются, тобто наближаються до кінців бівалента і спадають з нього. Таким чином, у міру наближення до метафази першого ділення число хіазм зменшується.
У метафазі I ділення мейозу район центромери кожної хромосоми сполучений (на відміну від метафази мітоза) ниткою веретена лише з одним полюсом клітки, причому центромери гомологів, що розійшлися, завжди пов'язані з протилежними полюсами.
Анафазе I ділення мейозу не передує розщеплювання центромери, як при мітозі, і тому до полюсів відходять не хроматиди, а цілі хромосоми, що складаються з двох хроматид. Проте, оскільки гомологічні хромосоми розходяться до різних полюсів, перше мейотичне ділення приводить до редукції числа хромосом. Іншими словами, по числу хромосом продукти I ділення мейозу стають гаплоїднимі. Проте у зв'язку з тим, що хромосоми в них зберігають подвійність, тобто містять дві хроматиди, кількість ДНК зменшується лише до 2с.
Друге ділення мейозу, наступне після короткого проміжку – інтеркінезу, приводить у відповідність число хромосом і вміст ДНК. Формально воно нагадує мітоз. На початку анафази відбувається розділення центромери, сестринські хроматиди стають дочірніми хромосомами і розходяться до полюсів. Таким чином, кожна з чотирьох клітин, що утворилися унаслідок двох мейотичних ділень однієї клітки, що пройшла предмейотичну s-фазу, міститиме п хромосом і з ДНК.
Отже, головна відмінність мейозу від мітоза – кон'югація гомологічних хромосом з подальшою розбіжністю їх в різні гамети. Точність розбіжності обумовлена точністю кон'югації, а остання – ідентичністю молекулярної структури ДНК гомологів.
2.3 Каріотип
Каріотипом називається хромосомний комплекс вигляду зі всіма його особливостями: числом хромосом, їх формою, наявністю видимих під світловим мікроскопом деталей будови окремих хромосом. Інколи термін «каріотип» вживають по відношенню до хромосомного набору одиничної клітини або групи тканинних клітин.

Число хромосом видоспецифічне. Хоча закономірності, що характеризують каріотип, інколи і відображають еволюцію певних видів, в цілому по структурі каріотипу прямо судити про систематичне положення виду не можна.
У більшості вищих тварин і рослин одна пара хромосом у особин однієї з родів гетероморфна. Ці несхожі хромосоми називаються статевими. Зокрема, у ссавців і в дрозофіли клітини чоловічих організмів мають Х- і y-хромосоми. В багатьох видів y-хромосома відсутня. Всі останні хромосоми називаються аутосомами.
Таким чином, завдяки дослідженням цитологів в кінці XIX – початку ХХ ст була обгрунтована роль ядра в спадковості, а спостереження за поведінкою хромосом в мітозе і мейозі привели до висновку, що саме з ними пов'язана передача спадкових ознак.
3. Секрети генетичного коду
У організмі кожної людини – своя спадкова конституція, характерна лише для нього. Саме з цим пов'язана тканинна несумісність, що виявляється, зокрема, при пересадці органів і тканин від одного організму іншому. «Чужа» шкіра, наприклад, зі своїми особливими молекулами вступає в небажані реакції з організмом «господаря». Вона викликає появу білків – антитіл – і в результаті не «приживається». Аналогічне явище спостерігається і при пересадці окремих органів.