Реферат: Механізми інактивації потенціал-залежних К+ каналів
Вступ.
При деполяризації мембрани багато потенціал-залежних йонних каналів, особливо деякі К+ канали переходять в довготривалий стан непровідності, чи інактивації. В результаті цього істотно змінюється транзієнтна проникність мембрани для калію, не зважаючи на тривалу і сильну деполяризацію. Така властивість деяких К+ каналів дозволяє дуже тонко регулювати палітру активності збудливих клітин через різну частоту потенціалів дії. Тому, вочевидь, дуже важливо для біологів та біомембранологів розуміти молекулярні механізми, які викликають інактивацію К+ каналів та можливі способи фізіологічної регуляції цього процесу.
Інактивація К + каналів
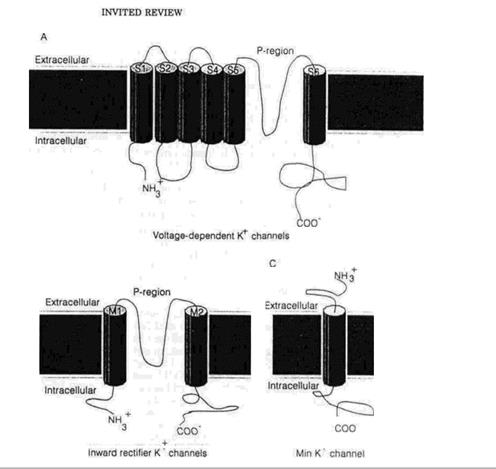 |
На відміну від гомогенної інактиваційної поведінки потенціал-залежних Na+ каналів, потенціал-залежні К+ канали, досліджені в їхньому природному середовищі, показують широку різноманітність інактиваційних часових інтервалів. Інактивація цих каналів спостерігається в діапазоні від кількох мілісекунд – швидка інактивація (як у А-струму, описаного в нейронах молюсків) до майже помітного процесу тривалістю кілька тисяч мілісекунд, (як у випадку струму затриманого випрямлення в гігантському аксоні кальмара). Така різноманітність також спостерігається в дослідах з клонованими К+ каналами, експресованими в гетерогенному середовищі. Наприклад, ShakerB канал інактивується з часовою константою 1-3 мс, тоді як відповідний канал drk1 демонструє значно повільнішу інактивацію. Так само, К+ канали, клоновані з мозку ссавців, показують дуже відмінні інактиваційні інтервали. В деяких випадках неструктурні білки, що утворюють minК+ канали (мал.1) не піддаються інактивації навіть після деполяризаційних імпульсів тривалістю кілька секунд. Ця різноманітність інактиваційних інтервалів навіть між дуже близькими між собою продуктами генів допомогла краще зрозуміти механізми інактивації К+ каналів.
NH 2 – кінцевий домен як інактиваційної частинки.
Відомо, що Shaker канали - це родина різних, але близько споріднених білків, які кодуються транскриптами альтернативного сплайсингу. Існує 5 альтернативно сплайсованих NH2 – кінцевих варіантів Shaker і 2 СООН – кінцевих. Після того, як вдалося встановити продукт Shaker локусу дрозофіли, дослідники припустили, що альтернативні транскрипти одного й того ж самого локусу індукують експресію К+ каналів з різною інактиваційною поведінкою.
Таким чином, наприклад, струм через Shaker А2 – канал, який відрізняється від ShakerD 2 лише своїм NH2 – кінцевим доменом, на 90% інактивується протягом кількох мілісекунд, тоді як струм через ShakerD 2 канал інактивується лише на 15% і значно повільніше.
IversonandRudy створювали конструкції з різними комбінаціямиNH2 – та СООН – кінцевих варіантів Shaker і помітили виникнення як інактиваційних, так і неінактиваційних струмів після їхньої експресії. Помічена різниця може бути віднесена на рахунок присутності саме NH2 – кінцевих доменів в кожній конструкції. Ці результати дозволили твердити про те, що структури, розташовані на NH2 – кінці (можливо це цитоплазматичні домени) можуть бути задіяні в інактиваційному процесі.
Працюючи з Shaker В каналами Hoshiта ін. простежили, що обробка трипсином внутрішнього боку збудливої мембрани катастрофічно сповільнює інактивацію ізольованих каналів, не впливаючи на процеси активації та проникності. Ці дані узгоджуються з припущенням про участь цитоплазматичного домену в процесі інактивації і через це вчені зосередили свою увагу на аналізі амінокислотної послідовності кінцевого NH2 домену Shaker В білку. Hoshiта співробітники робили систематичні делеції амінокислот і встановили, що делеції, які включають перші 22 амінокислоти змінюють швидкий інактиваційний процес на зразок як це робить трипсин. Такі делеції збільшують середній час, протягом якого канали перебувають у відкритому стані, збільшують кількість відкритих протягом серії каналів та зменшують тривалість інтервалів між серіями. Ці дані відповідають результатам про сповільнення інактивації макроскопічних струмів. Делеції поза ділянкою перших 22 амінокислот не порушують інактивацію, але зменшують середню тривалість відкритості каналів пропорційно до довжини делетованої ділянки. Ці спостереження доводять роль перших 22 амінокислот як інактиваційної “кульки”, а решти NH2 кінцевого домену як “ланцюжка”, що сполучає кульку з каналом.
Прогноз про те, що, виходячи з моделі “кульки і ланцюжка” додавання ізольованих кульок до цитоплазматичного боку каналів, не здатних до інактивації відновить цю їхню здатність підтвердився експериментами Zagottaetal. В цих експериментах рівень відновленої каналами інактивації залежав від концентрації доданого кулькового пептиду. Ефект дії пептиду виявився зворотнім і не спостерігався при попередній обробці пептиду трипсином.
Shaker В інактиваційна кулька має первинну структуру, що містить 11 гідрофобних або незаряджених залишків та наступну групу з восьми сильно полярних амінокислот. Aldrichта співробітники більш детально дослідили внесок в інактивацію кожної частини кулькового домену. Вони зробили серії селективних мутацій і довели важливість як гідрофобної, так і зарядженої частини домену для процесу інактивації. Модель Shaker В інактиваційної кульки може бути застосована і до інших типів потенціал-залежнних каналів. Наприклад, було показано, що делеція перших 25 амінокислот NH2 кінцевого домену серцевих К+ каналів щурів призводить до значного зниження швидкої інактивації. Також делеція NH2 кінцевого домену (амінокислот 1-110) RCK4 призводить до експресії повільно інактивованого струму, на відміну від А-типу струму, який виникає внаслідок екпресії RCK4 транскрипту повної довжини. Найбільш визначним є той факт, що Shaker В кульковий пептид також має здатність блокувати (інактивувати?) еволюційно більш віддалені К+ канали. Було показано, що інші потенціал-залежні Са-активовані калієві канали, і ц-АМФ-залежні канали блокуються Shaker В кульковим пептидом, якщо його додавати внутрішньоклітинно. Ці спостереження особливо помітні через те, що такі канали за нормальних умов не виявляють інактивації. Дані, отримані з Shaker К+ каналами так само як і з maxi К+ струмами пролили світло на тонкі процеси взаємодії між інактиваційною кулькою та каналом.
Взаємодія кулькового пептиду та рецептора.
Ідея про те, що як гідрофобний так і заряджений кінець кульки роблять свій внесок у взаємодію між кульковим пептидом і рецептором більш інтенсивно досліджувалась шляхом спострежння ефекту мутантних пептидів на інактивацію Shaker В і maxi К+ . В обох випадках було продемонстровано, що зміна загального заряду кульки (внаслідок мутації амінокислот в СООН ділянці) впливає на рівень інактивації як макроструму так і струму через ізольовані окремі канали. Цікаво, що ці зміни заряду пептиду модулюють лише рівень включення інактивації, в той час як рівень виключення інактивації залишається в основному незмінним. Таким чином, збільшення позитивного заряду кульки підвищує рівень включення, і навпаки, зниження позитивного заряду або підвищення негативного зменшує рівень включення інактивації. Ці ефекти електричного заряду максимальні в умовах низької іонної сили і мінімальні при високих сольових концентраціях. Всі разом ці результати було інтерпретовано з точки зору різноманітних елктростатичних взаємодій між позитивно зарядженою кулькою і негативно зарядженою поверхнею в, або поблизу внутрішнього гирла каналу. Електростатичний потенціал, індукований цими негативними зарядами буде спрямовувати кульку до її рецептора. З іншого боку заміна гідрофобних амінокислот полярними і незарядженими залишками з NH2 – кінця кулькового пептиду зменшує рівень макроскопічної інактивації; і цей ефект спостерігається головним чином через збільшення неблокованого рівня. Особливо важливими є мутації, які змінюють гідрофобність в позиції L7 (заміна лейцину). З цих даних видно, що тоді, коли інактиваційний домен взаємодіє зі своїм рецептором, гідрофобні взаємодії зв’язують їх докупи і сила цих взаємодій визначає рівень при якому канали повертаються до неінактивованої конформації, точніше до рівня, коли кулька дисоціює з рецептора.
Однак, профіль гідрофобності консервативний, що виходить з припущення про те, що гідрофобні взаємодії визначають стабільність комплексу кулькового пептиду з рецептором. На підтвердження цієї думки, було показано, що цитоплазматичний домен (коли він приєднується в якості химерної частинки) зв’язує сегмент 3 або 4 (мал.1) потенціал-залежних Na-каналів також здатний діяти як інактиваційна частинка для калієвих каналів. 3-4 зв’язок було запропоновано як інактиваційну кульку для Na-каналів. З цих досліджень і було зроблено висновок про те, що рецептор кулькового пептиду може мати негативний поверхневий потенціал, як було попередньо показано для внутрішніх ворітних калієвих струмів, і також обов’язково має мати гідрофобну поверхню, яка здатна взаємодіяти з першою половиною кулькового пептиду. Хоча реакція між кульковим пептидом і ShB D 6-46 К+ каналом бімолекулярна, в проміжку часу, який відповідає відновленню від заблокованого стану можна розрізнити два різні стани блоку. Дуже складно пояснити молекулярну природу кожного стану, але з огляду на дуже високий рівень константи зв'язування Q10 в реакції пептиду з каналом, Murrell-LagnagoandAldrich запропонували гіпотезу про те, що два стани виникають через присутність кількох різних структурних форм пептиду в розчині. Підвищення температури збільшує концентрацію пептиду з правильною структурою для зв'язування.
Isacoffetal. встановили, що мутації в цитоплазматичній петлі, яка приєднує трансмембранні домени S4 і S5 спричинюють дикий тип інактивації в ShakerB і drk 1 , в яких інактивація індукується додаванням ShakerB - кулькових пептидів з внутрішньої сторони плазматичної мембрани. Відповідаючи дослідженням з кульковим пептидом, гідрофобний залишок має особливе значення для стабілізації інактивованого стану. Мутації цієї ділянки також змінюють властивості йонної провідності, з чого можна припустити їхній близький зв'язок з канальними структурами, через які перебігає йонний струм.
Деякі автори пропонують вважати, що близькість залишків, задіяних в процесі швидкої інактивації до S4 домену може створювати основу тісного зв'язку між активацією та інактивацією. Конформаційні зміни внаслідок вилучення заряджених залишків S4 спіралі можуть збільшувати афінність кульки до її рецептора. На підтримку взаємодії між доменами, задіяними в процесах активації та інактивації, Bozanillaпоказав, що присутність інактиваційної частинкиShakerB каналів сповільнює переміщення воротних зарядів під час припинення деполяризаційних пульсів. Ці результати змушують думати про можливість конформаційних перебудов, а не просто взаємодію кульки з рецепторами в процесах активації та інактивації.
Механізм блокування кульковим пептидом.
Як зазначалося вище, інактиваційна частинка поводиться схоже на блокатор йонного струму. Це було продемонстровано, використовуючи ТЕА блокатори К+ каналів в гігантському аксоні кальмара. Для того, щоб зрозуміти механізм блокування інактиваційною частинкою, необхідно повернутися до класичної біофізики.
Armstrong встановив, що внутрішньоклітинний ТЕА блокує лише канали у відкритому стані, що пов'язується з думкою про те, що ТЕА глибоко проникає в середину каналів, перекриваючи потік йонів. Така ідея протилежна до думки про можливу алостеричну дію, яку виявляє ТЕА, індукуючи довготривалі конформаційні зміни і порушення провідності йонів через канальні структури.
Виходячи з припущення про локалізацію блокатор-зв'язуючої ділянки, можна прогнозувати, що збільшення йонного струму через канал здатне дестабілізувати взаємодію блокатора з каналами і таким чином пом'якшувати блокаду.
Властивості інактиваційної частинки ShakerB як блокатора відкритих каналів також були продемонстровані.
DemoandYellen показали, що час відновлення від інактивації (тобто час, потрібний для того, щоб канальний білок повернувся від інактивованого стану до закритого неінактивованого стану) скорочується за присутності високих концентрацій К+ в зовнішньому середовищі. Це узгоджується з ідеєю про "пом'якшення інактивації".
Експерименти з ізольованими каналами підтвердили точку зору про те, що канали інактивуються у відкритому стані. Точніше, для того, щоб інактивуватись, канал спочатку має відкритися, а після приєднання, інактиваційна частинка заважає каналові повернутися до закритої конформації. Отже, просту кінетичну модель актиації можна зобразити таким чиом: С-О-І. Для того, щоб перейти від інактивованого до закритого стану, каналові необхідно відкритися. (мал.2) Інактиваційний ShakerB кульковий пептид поводиться як блокатор відкритих каналів.
Стехіометрія інактиваційної реакції.
З вищесказаного випливає очевидність того, що потенціал-залежні канали - це тетрамери, утворені ідентичними чи змішано - незалежними субодиницями. Коли це так, то кожен канал має 4 незалежні NH2 - кінці, потенційно здатні бути інактиваційними частинами. Скільки їх потрібно, щоб відбулася інактивація?
MacKinnonetal. підійшли до цього питання створюючи субодиничні форми Shaker , в яких присутність інктиваційної кульки була пов'язана з мутацією, що робить канал стійким до блокади СТХ, але не має відношення до інактивації. Цю форму каналу ін'єктували в ооцити Xenopuslaevisразом з токсин-нечутливими неінактиваційними субодиницями, і очікувалась експресія тетрамерів з хаотичним випадковим складом. Достатньо лише одного токсин-зв'язуючого сайту, щоб підтвердити чутливість до токсину, коли лише однієї кульки достатньо для інактивації каналу, тоді всі токсин-чутливі канали з вбудованими 1-4 токсин чутливими субодиницями, які несуть кульки, мають інактивуватися.
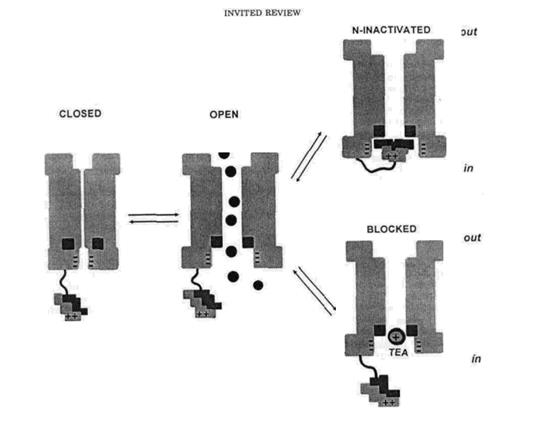 |
Насправді, всі швидко інактиваційні компоненти, індуковані ін'єкцією змішаних субодиниць, блокувалися СТХ. Однак інактивація каналів, які містили менше ніж 4 інактиваційні частинки, була повільнішою ніж у тих, які містили всі 4 кульки. Такі дані можна трактувати аналогічно до доза-залежного рівня інактивації ShB D 6-46 каналів, на які діяли вільним кульковим пептидом, при чому зростання інактиваційного рівня відбувалося коли пептид був більш позитивно зарядженим. В будь-якому разі припускається, що збільшення концентрації кулькового пептиду біля його рецептора, збільшує вірогідність їхнього зв'язку і відтак - рівня інактивації; тим не менш, лише одна кулька взаємодіє з гирлом каналу.
b - субодиниця швидкої інктивації.
Більшість досліджень молекулярного та біофізичного механізмів швидкої інактивації К+ каналів зосереджувалось навколо NH2 кінцевого домену а- субодиниці канального білку. Однак, деякими авторами було також висловлено думку про те, що білкові домени з інших субодиниць можуть спричинювати швидку інактивацію неінактивованих каналів, коли їх експресувати разом з а- субодиницею.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--