Реферат: Молекулярные основы эволюции дифференцировки развития и старения
- В результате заполнения бреши и лигирования происходит дупликация сайта-мишени.
- Таким способом может осуществляться нерепликативная транспозиция Тn 10.
- Эту реакцию катализирует транспозаза Тn 10, которая ускоряет разрезание на концах Тn 10, а также, по-видимому, воссоединение разорванных концов.
- При репликативной транспозиции общий промежуточный продукт претерпевает совсем другие превращения.
- интермедиат по своей структуре аналогичен кольцевой ДНК с двумя репликативными вилками.
- Если репликация начинается в каждой вилке, то конечный продукт представляет собой коинтеграт-кольцо, содержащее и донорную, и реципиентную ДНК, а также копии транспозирующегося элемента, реплицированные полуконсервативным путем.
- При разрешении коинтеграта в конце концов осуществляется транспозиция новой копии мобильного элемента в другой сайт-мишень. Разрешение может происходить путем гомологичной рекомбинации между двумя копиями транспозирующегося элемента с участием рекомбинационного аппарата Е.соli. (прозрачка 6).
Генетическая изменчивость бактерий при транспозиции
Мобильные элементы вызывают генетическую нестабильность поблизости от участка своей локализации, особенно в процессе репликативного механизма транспозиций.
В зависимости от того, как внесены разрывы в ДНК-мишень, получится либо делеция (Выпадение участка хромосомы из ее внутренней области), либо инверсия (перестановка) генетического материала между местом расположения транспозона и мишенью его перемещения. В связи с этим интересно отметить, что хромосомы родственных видов бактерий отличаются друг от друга многочисленными перестройками именно этого типа. По-видимому, мобильные элементы сыграли существенную роль в дивергенции и видообразовании бактерий.
1) Встраивание IS‑элементов поблизости от молчащего гена может приводить к его активации за счет транскрипции с промотора IS– элемента, т.е. изменяется регуляция бактериального гена (пр. 8а).
2) Очень важно, что мобильные элементы служат подвижными участками гомологии, гомологическая рекомбинация между которыми может приводить к дупликациям генов (рис. 76 Коничев). Считается, что дупликация – один из основных путей эволюционного возникновения новых функций. Действительно, «лишняя» копия гена выходит из-под давления естественного отбора и получает возможность накапливать изменения. Чаще всего это приведет к утрате какой бы то ни было функции, но иногда может получиться ген с новыми функциями.
3) Нельзя забывать и тот факт, что клетка может получить селективное преимущество за счет приобретения в составе транспозона гена, который сам по себе способен оказаться выгодным для бактерии в определенных условиях. Действительно, на транспозонах «путешествуют» гены устойчивости к различным бактериальным ядам, в том числе к тяжелым металлам и антибиотикам, гены дополнительных метаболических путей, позволяющие использовать необычный источник углерода, наконец, гены некоторых токсинов, делающие бактерии патогенными и позволяющие им тем самым существенно изменить образ жизни. Сказанное в равной степени относится и к плазмидам, поскольку большинство полезных для клетки-хозяина плазмидных генов находится в составе транспозонов.
4) Если элемент встроен в один из генов полицистронного оперона, то встроенный элемент может влиять на экспрессию последующих генов, либо останавливая транскрипцию в сайте терминации, находящемся внутри его самого (прозрачка 8,6), либо подавляя трансляцию дистальных кодирующих последовательностей мРНК (прозрачка 8, в)
5) Когда один и тот же геном служит и донором, и реципиентом при образовании коинтеграта (внутримолекулярная транспозиция), способность мобильных элементов вызывать перестройки в соседних участках ДНК приводит к другим мутационным эффектам. Все происходящие при этом события, включая делеции и инверсии, связаны с соединением одного или двух концов мобильного элемента с новыми последовательностями ДНК (рис. 10.13); детали этих процессов до конца не установлены.
Подвижные генетические элементы у эукариот
Существенную часть генома эукариот (10–30%) составляют повторяющиеся последовательности, имеющие определенную структурную организацию и способные перемещаться в геноме как в пределах одной хромосомы, так и между хромосомами. Они получили название подвижных генетических элементов. Элементы включают в свой состав от тысячи до десятков тысяч нуклеотидных пар.
Наибольшее количество подвижных элементов обнаружено в геноме растений (до 50%). Подвижные генетические элементы обычно рассеяны по геному, но могут концентрироваться в отдельных участках хромосом.
Виды мобильных элементов эукариот
Различают два (по признаку молекулярных механизмов перемещения) основных класса подвижных генетических элементов:
1. Транспозоны
Эти элементы ограничены инвертированными повторами (последовательностями, направленными навстречу друг другу), как и некоторые транспозоны прокариот. Примерами их могут служить Р-элемент дрозофилы и Ас-элемент кукурузы (рис. 92 Коничев). В геномах этих организмов насчитывается по 30 – 50 копий таких элементов. Полные копии (часть копий дефектна, так как имеет внутренние делеции) содержат открытые рамки считывания, кодирующие транспозазу (рис. 92).
Схема перемещения транспозона показана на рис. 93. Инвертированные повторы необходимы для перемещения элемента, которое осуществляется благодаря их сближению друг с другом и узнаванию транспозазами. Инвертированные повторы сближаются и точно отрезаются от соседних участков ДНК хозяина. Вырезанный транспозон внедряется в район вносимого транспозазой разрыва в молекуле-мишени и сшивается с ДНК хозяина в новом месте. Разрыв и зашивание осуществляются транспозазой и вспомогательными белками. Транспозаза может кодироваться как самим подвижным элементом, который будет перемещаться, так и другой копией элемента, локализованной в том же геноме в отдалении. Р-элементы обычно встраиваются в определенные сайты, консенсусная последовательность которых такова: 5'-ГГЦЦАГАГ При встраивании Р-элемента этот сайт-мишень дуплицируется.
![]()
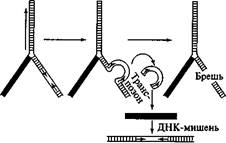
Встраивание
Рис. 93 Перемещение транспозона
Брешь в ДНК после вырезания транспозона может заделываться либо с помощью гомологичного отрезка ДНК, либо за счет сшивания разорванных концов (это особенно характерно для транспозонов растений).
Эффект от перемещения транспозона эукариот