Реферат: Рекомбинантные вакцины (Генная инженерия)
Расщепление полирибонуклеотидной цепи РНК может осуществляться в присутствии различных ферментов. Для этой цели используют ферменты: рибонуклеазу H, нуклеазы, ковалентно связанные с олигонуклеотидами (например, стафилококковая нуклеаза), рибозимы (например, рибозим L-19).
Исторически первым способом направленной ферментативной фрагментации РНК (называемой также адресованной, сайт-направленной или сайт-специфической фрагментацией РНК) является разработанный в Московском университете метод гидролиза РНК ферментом РНКазой H в присутствии комплементарных олигодезоксирибонуклеотидов. Этот метод до сих пор остается наиболее универсальным способом направленной фрагментации РНК.
Метод основан на свойстве РНКазы Н расщеплять полирибонуклеотидную цепь РНК в составе ДНК-РНК-гетеродуплекса. К участку РНК, по которому планируется провести ее фрагментацию, синтезируется комплементарный олигодезоксирибонуклеотид длиной в 6-10 нуклеотидных остатков. Далее получают комплекс этого олигонуклеотида с РНК, который затем обрабатывают ферментом. В работе обычно используют РНКазу Н из Escherichia coli - вполне доступный фермент, который может быть довольно легко очищен от всех сопутствующих нуклеазных примесей. Этот метод нашел достаточно широкое применение для сайт-специфического расщепления вирусных и рибосомных РНК, а также для идентификации продуктов процессинга некоторых РНК.
Важным достоинством рассматриваемого метода является и то, что в результате гидролиза РНК рибонуклеазой Н образуются фрагменты, у одного из которых на 5'-конце содержится фосфатная группа, а у другого 3'-конец свободен (нефосфорилирован). Такие фрагменты можно прямо использовать в реакции ферментативного лигирования.
Серьезным ограничением этого метода является то, что участок РНК, с которым связывается комплементарный ему олигодезоксирибонуклеотид, должен иметь однотяжевую конформацию и находиться на поверхности макромолекулы РНК, представляющей собой в условиях расщепления компактную глобулу с развитой вторичной структурой. В отдельных случаях это ограничение удается преодолеть, используя достаточно длинные олигодезоксирибонуклеотиды (15-20-членные), которые предварительно отжигают с частично или полностью денатурированной РНК.
Другое ограничение метода фрагментации полирибонуклеотидов РНКазой Н в присутствии комплементарных олигодезоксирибонуклеотидов заключается в том, что в общем случае предсказать, какая из фосфодиэфирных связей в гетеродуплексе (или в непосредственной близости от него) подвергнется расщеплению, не удается. Более того, фермент зачастую гидролизует не одну, а несколько соседних межнуклеотидных связей. Ясно, что для последующего конструирования рекомбинантных РНК такие фрагменты могут оказаться непригодными.
Расщепление РНК может проводиться и с участием нуклеаз, ковалентно связанных с олигонуклеотидами. Идея, лежащая в основе данного подхода к направленной фрагментации РНК, восходит к концу 60-х годов, когда Н. И. Гриневой и ее сотрудниками был предложен метод олигонуклеотид-направляемой модификации нуклеиновых кислот. Принцип этого метода заключается в том, что с 5'- или 3'-концевым остатком олигонуклеотида, комплементарного заданному району ДНК и РНК, связывают модифицирующий агент, который после образования дуплекса атакует одно из ближайших к нему оснований. Реагентами этого типа удалось направленно фрагментировать фенилаланиновую тРНК из дрожжей, РНК-компонент (M1 РНК) РНКазы Р, а также 16S рибосомную РНК E .coli .
Однако, как и в случае РНКазы Н, расщепление РНК олигонуклеотид-нуклеазой зачастую проходит по нескольким межнуклеотидным связям, что несколько ограничивает возможности применения этого метода для получения рекомбинантных РНК.
Для расщепления РНК применяют также рибозимы (природные РНК и синтетические полирибонуклеотиды, способные катализировать целый ряд превращений у других РНК). Первый рибозим, напоминающий по своим свойствам эндонуклеазы рестрикции, был получен Т. Чеком. В научной литературе его обозначают как рибозим L-19. Этот рибозим представляет собой РНК длиной в 395 нуклеотидных остатков, в 5'-концевой области которой имеется гексануклеотидная последовательность GGAGGG, ответственная за специфичность расщепления предшественника 26S РНК при самосплайсинге. Эта последовательность комплементарна CUCUCU последовательности, расположенной на 3'-конце первого экзонного участка предшественника 26S РНК. Если эту РНК заменить другой, но обязательно содержащей доступную для комплементарного связывания CUCUCUA последовательность, то рибозим L-19 в присутствии гуанозина или гуаниловых нуклеотидов со свободной 3'-гидроксильной группой расщепит ее.
Источником необходимых участков РНК может служить такой способ, как синтез фрагментов РНК. Ферментативный синтез сегментов РНК осуществляют с использованием разнообразных генно-инженерных конструкций. Однако в настоящее время в подавляющем большинстве случаев для препаративного получения РНК используются РНК-полимеразы, закодированные в геномах ряда ДНК-содержащих бактериофагов (Т3, Т7 и SP6). Они характеризуются очень высокой активностью и, в отличие от клеточных РНК-полимераз, состоят из одной полипептидной цепи. Важно также то, что инициация и терминация синтеза РНК этими полимеразами происходит на одном определенном нуклеотидном остатке.
Полученные одним из способов фрагменты нуклеиновых кислот в дальнейшем встраивают в векторные молекулы.
Подводя итог изложенному, можно сделать вывод, что получение генов протективных белков возбудителей представляет собой достаточно сложную задачу. Однако ее решение необходимо для создания в конечном счете рекомбинантных противовирусных вакцинных препаратов.
1.2. ВЫБОР ВЫСОКОАКТИВНОЙ И ХОРОШО ИЗУЧЕННОЙ В
ИММУНОЛОГИЧЕСКОМ ОТНОШЕНИИ МОДЕЛИ ВЕКТОРА-НОСИТЕЛЯ И КЛОНИРОВАНИЕ СООТВЕТСТВУЮЩЕГО ГЕНА
1.2.1. ПОЛУЧЕНИЕ РЕКОМБИНАНТНЫХ ДНК
Суть конструирования рекомбинантных ДНК заключается во встраивании фрагментов ДНК, среди которых находится интересующий нас участок ДНК, в так называемые векторные молекулы ДНК (или просто векторы) - плазмидные или вирусные ДНК, которые могут быть перенесены в клетки про- или эукариот и там автономно репли-цироваться. На следующем этапе проводится отбор тех клеток, которые несут в себе рекомбинантные ДНК (с помощью маркерных признаков, которыми обладает сам вектор), и затем индивидуальных клонов с интересующим нас сегментом ДНК (используя признаки или пробы, специфичные для данного гена или участка ДНК).
При решении ряда научных и биотехнологических задач конструирование рекомбинантных ДНК требует также создания систем, в которых обеспечивается максимальная экспрессия клонируемого гена.
Существует три основных способа встраивания чужеродной ДНК в векторные молекулы. В первом случае 3'-концы фрагментов ДНК, среди которых находится интересующий нас участок ДНК (ген или его сегмент, регуляторный район), с помощью фермента терминальной нуклеотидилтрансферазы наращиваются гомополинуклеотидной последовательностью (например, поли (Т)). 3'-концы линейной формы векторной ДНК тем же способом наращиваются комплементарной ей гомополинуклеотидной последовательностью (то есть поли (А)). Это позволяет соединить две молекулы ДНК путем комплементарного спаривания искусственно полученных "липких" концов.
Во втором случае "липкие" концы создаются с помощью расщепления молекул ДНК (как векторной, так и содержащей интересующий нас фрагмент) одной из эндонуклеаз рестрикции (рестриктаз). Рестриктазы характеризуются исключительно высокой специфичностью. Они "узнают" в ДНК последовательность из нескольких нуклеотидных остатков и расщепляют в них строго определенные межнуклеотидные связи. Поэтому даже в ДНК больших размеров рестриктазы вносят ограниченное число разрывов.
Третий способ представляет собой комбинацию двух первых, когда липкие концы ДНК, образованные рестриктазой, удлиняются синтетическими последовательностями (рис. 1).
Концы фрагментов ДНК можно превратить в "липкие", наращивая их двутяжевыми олигонуклеотидами ("линкерами"), в состав которых входит участок узнавания рестрикта-
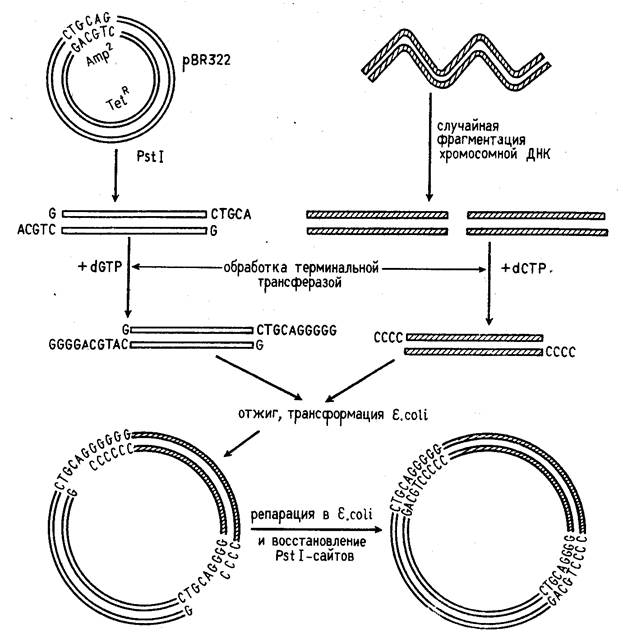
Рисунок 1. Схема конструирования рекомбинантной ДНК с помощью рестриктаз PstI и поли(G)- поли(С)-линкера.
зой. Обработка такого фрагмента данной рестриктазой делает его пригодным для встраивания в векторную молекулу ДНК, расщепленную той же рестриктаэой. Часто в качестве "линкера" применяются полинуклеотидные фрагменты, которые содержат специфические участки сразу для нескольких рестриктаз (их называют "полилинкерами").
После встраивания чужеродной ДНК в вектор их ковалентное сшивание осуществляется ДНК-лигазой. Если же размер бреши в рекомбинированной молекуле превышает одну фосфодиэфирную связь, она застраивается in vitro с помощью ДНК-полимеразы или in vivo с помощью репарирующих систем клетки.
1.2.2. ПОЛУЧЕНИЕ РЕКОМБИНАНТНЫХ РНК
Получение рекомбинантных РНК обычно осуществляют методами ферментативного или химического лигирования РНК. Кроме того, недавно появилась принципиально новая возможность встраивания сегмента РНК в заданное положение других молекул РНК с помощью рибозимов.
Ковалентное сшивание отдельных сегментов РНК при получении рекомбинантных молекул, как правило, осуществляют с помощью Т4 РНК-лигазы. Т4 РНК-лигаза закодирована в геноме бактериофага Т4. Ее выделяют из клеток E .coli , зараженных этим фагом. Фермент сшивает друг с другом однотяжевые олиго- и полирибонуклеотиды. Для работы Т4 РНК-лигазы необходим источник энергии - аденозинтрифосфат. На рис. 2 приведена схема ферментативного лигирования двух коротких олигонуклеотидов. Как видно из этой схемы, акцептором в реакции лигирования служит полностью дефосфорилированный, а донором - полностью фосфорилированный по концевым нуклеотидным остаткам олигонуклеотид. Это предотвращает возможность сшивания однотипных олигонуклеотидов.
Эффективность ферментативного лигирования достаточно длинных полирибонуклеотидов сильно варьирует и ее трудно предсказать исходя только из нуклеотидной последовательности сегментов РНК. Наилучшие результаты получены в тех случаях, когда сшиваемые концы полирибонуклеотидов были пространственно сближены за счет комплементарного связывания соседних с ними участков РНК.
Недавно было установлено, что протяженные сегменты РНК (длиной в 200-300 остатков) могут быть с высоким выходом сшиты Т4 ДНК-лигазой. При этом "стыковка" сегментов осуществляется с помощью олигодезоксирибонуклеотида, комплементарного 3'-концу одного сегмента и 5'-концу другого.
Метод химического лигирования основан на активации концевой фосфатной группы одного из двух сшиваемых сегментов РНК водорастворимым карбодиимидом или
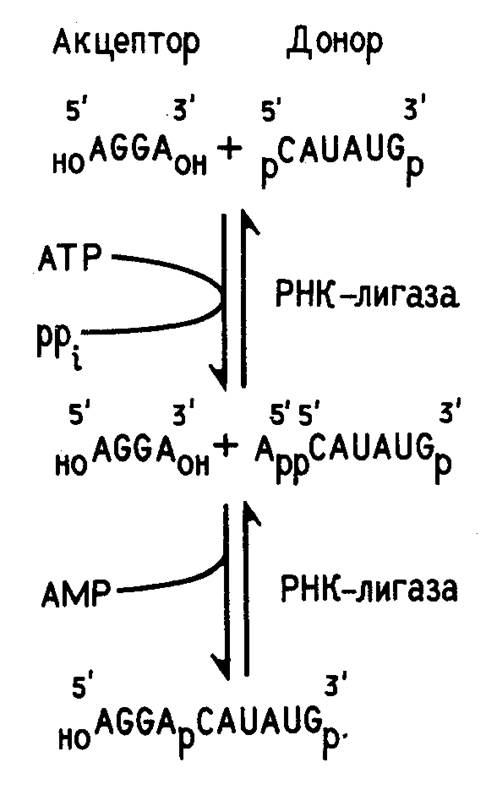
Рисунок 2. Схема сшивания двух олигорибонуклеотидов с помощью Т4 РНК-лигазы.
BrCN. В случае BrCN реакция протекает очень быстро и не сопровождается модификацией нуклеотидных остатков, хотя под действием карбодиимидов фосфодиэфирная связь образуется с более высоким выходом. Для того, чтобы обеспечить сближенность сшиваемых концевых нуклеотидных остатков в фрагментах РНК, было предложено использовать олигодезоксирибонуклеотиды, комплементарные обоим фрагментам в месте их стыка.