Реферат: Симбиоз
Среди симбиозов, образованных с участием водоросле й, наибольший интерес пре дставляет симбиоз водорослей с грибами, известный под названи ем лишайн и кового сим био за.
ВЗАИМООТНОШЕНИЯ ГРИБА И ВОДОРОСЛИ В ТЕЛЕ ЛИШАЙНИКА
Вопрос взаимоотношения гриба и водоросли в слоевище лишайника занимал умы ученых еще в конце прошлого столетия, да и в паше время продолжает волновать лихенологов. Со дня открытия С. Швендснера прошло более 100 лет. За этот период появилось не менее десятка теорий, пытающихся объяснить отношения между грибом и водорослью, однако среди них нет ни одной общепризнанной и окончательно доказанной. С. Ш в е н д е не р, обнаружив, что лишайник состоит из гриба и водоросли, предположил, что гриб в слоевище паразитирует на водоросли. Однако он ошибочно отвел грибу роль хозяина, а водоросли — раба.
Но уже в те времена некоторые ученые выдвинули мысль о двустороннем паразитизме компонентов лишайника — гриба на водоросли и водоросли на грибе. При этом было высказано предположение, что гриб и водоросль в слоевище лишайника находятся в полном морфофизиологическом единстве и связаны между собой так же, как корни и листья цветковых растений. Такое сравнение, безусловно, было совсем необоснованным.
Наибольшее распространение среди ученых того времени получила теория мутуалистического симбиоза. Сторонники этой теории считали, что в слоевище лишайника гриб и водоросль находятся во взаимовыгодном симбиозе: водоросль “снабжает” гриб органическими веществами, а гриб “защищает” водоросль от чрезмерного нагревания и освещения и “обеспечивает” ее водой и неорганическими солями. Однако в 1873 г. этой идеалистической теории был нанесен удар. Известный французский исследователь Е.Борн е, изучая анатомическое строение слоевища лишайников, обнаружил внутри водорослевых клеток грибные отростки — гаустории, всасывающие органы гриба. Это позволяло думать, что гриб использует содержимое клеток водорослей, т. е. ведет себя как паразит.
За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных форм абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или проникают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, грибному компоненту.
О том, что в слоевище лишайника происходит обмен веществами между грибом и водорослью, ученые стали говорить сразу после открытия двойственной природы лишайников. Однако некоторые экспериментальные подтверждения этим предположениям были получены лишь за последние три десятилетия. Применение новейших методов физиологических исследований с использованием меченых атомов углерода и азота, особых красящих веществ и некоторых других позволило установить, что гриб получает вещества, ассимилируемые водорослью, и ведет себя в слоевище лишайника как паразитический организм. Однако для существования как самого гриба, так и лишайника в целом необходимо, чтобы водоросль, окруженная со всех сторон грибными гифами, все-таки могла жить и более или менее нормально развиваться. Если гриб начнет проявлять себя слишком активно, поражать все без исключения водоросли и, использовав их содержимое, уничтожать их, это в конце концов может привести к гибели всех водорослей слоевища. Но тогда, уничтожив весь свой запас питания, по-
гибнет и сам гриб, а значит, перестанет существовать и лишайник.
Гриб должен использовать лишь часть водорослей, оставляя резерв — здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Учеными были замечены любопытные защитные реакции со стороны лишайниковых водорослей. Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.
Способность водорослей нормально развиваться и даже размножаться в слоевище лишайника сохраняется скорее благодаря умеренности паразитизма самого гриба.
Ученые отмечают, что степень паразитизма гриба на водоросли различна не только у разных видов лишайников, но даже в одном и том же слоевище. Резкий паразитизм обнаружен лишь у примитивных лишайников. Гаустории, проникающие глубоко внутрь протопласта водоросли, пока что были найдены лишь у наиболее просто организованных форм, в слоевище которых еще нельзя различить оформленных дифференцированных слоев. В слоевищах более высокоорганизованных лишайников часть клеток водорослей поражена грибными гифами, а остальные продолжают нормально жить и развиваться. Обычно у высокоорганизованных форм лишайников паразитизм гриба на водоросли носит весьма умеренный характер: прежде чем гриб убьет пораженные им клетки, успевает вырасти одно или несколько поколений водорослей.
Отношения между мико- и фикобионтом в слоевище лишайника не сводятся только к паразитизму гриба на водоросли. Ученые предполагают, что эти отношения гораздо сложнее. Еще в начале нашего века крупнейший русский лихенолог А. А. Е ленки н, изучая анатомическое строение лишайников, обнаружил в их слоевище некральные зоны водорослей — скопления отмерших, потерявших зеленую окраску клеток, расположенные несколько ниже зоны живых водорослей. К этим бесцветным мертвымклеткам водорослей тоже тянулись грибные гифы. Это привело А. А. Еленкина к мысли, что гриб в слоевище лишайника вначале проявляет себя как паразитический организм, поражая живые клетки водоросли и используя их содержимое. Затем, убив водоросль, гриб переходит к сапрофитному способу питания, поглощая и ее мертвые остатки. Таким образом, гриб в слоевище лишайника ведет себя и как паразит, и как сапрофит. И отношения между грибом и водорослью в слоевище лишайника А. А. Еленкин назвал эндопаразитосапрофитизмом.
Интересную мысль о взаимоотношении компонентов в слоевище лишайника высказал в 60-х годах нашего столетия крупнейший советский лихенолог А. Н. О к сне р. Он пришел к выводу, что водоросль в слоевище лишайника, полностью изолированная от внешней среды грибной тканью, обязательно должна забирать у грибного компонента все необходимые для своего существования вещества, за исключением тех органических соединений, которые она сама вырабатывает на свету в процессе ассимиляции углекислоты. К этим жизненно необходимым для водоросли веществам относится прежде всего вода, а также минеральные соли, азотистые и некоторые другие неорганические соединения. Следовательно, и водоросль в слоевище лишайника проявляет себя как паразит. Причем это вовсе не противоречит общему характеру ее питания. Как показало изучение лишайниковых водорослей в чистых культурах, многие из них, будучи большей частью автотрофными организмами, способны и к миксотрофному питанию.
Таким образом, ученые считают, что водорослевый и грибной компоненты лишайника находятся в очень сложных взаимоотношениях. Микобионт ведет себя как паразит и сапрофит на теле водоросли, а фикобионт, в свою очередь, паразитирует на лишайниковом грибе. При этом паразитизм фикобионта всегда носит более умеренный характер, чем паразитизм гриба.
 |
?????? ??? ??????????? ?? ????? ?????? ????? ?????? ?? ??? ??? '???????? ???? ?????????? ? ??????? ?????? ?? ???????????? ?????????????????: ????????? ????????? ????? ???????? ???????? ??? ??????????????? ?????????????. ?????? ???? ?? ????????? ?????????? ? ???????????? ? ????? ????????? ???????? ?????????? ? ????????????? ????????. ??? ??????? ????? ?????? ? ??????????, ??????? ? ????? ????????? ??????????? ? ??????? (??????????? ????????? ???????????? ??????????!), ????? ?? ??????? ????????????? ? ????????????? ????????. ????????, ??? ???????? ??????????? ? ??????????? ???? ??????? ????? ?????????? ? ???????? ?????? ????????. ????? ?? ??????? ?????????? ????????? ?? ??????? ?????? ??????????? ????????? ? ???????? ???????????, ?? ???? ??? ????????? ???????? ? ?? ?????? ??????????.
Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники — многолетние растения. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20—50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2—0,3 мм.
Кустистые и листоватые лишайники растут несколько быстрее — в год их слоевище увеличивается на 2—3 мм. Поэтому, чтобы вырастить взрослый лишайник в лаборатории, требуется не менее 20 лет, а может быть, и вся жизнь исследователя. Трудно проводить столь долговременный эксперимент!
Вот почему физиологические особенности лишайников, в том числе взаимоотношения компонентов, как правило, изучают на культурах изолированных мико- и фикобионтов. Этот метод очень перспективен, так как позволяет ставить длительные и воспроизводимые опыты. Но, к сожалению, данные, полученные этим методом, не могут полностью отразить те процессы, которые происходят в целом слоевище лишайника.
И тем более мы не вправе считать, что в природе, в естественных условиях, в слоевищах лишайника эти процессы протекают точно так же, как в культурах изолированных симбионтов. Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов лишайников, остаются пока лишь догадками.
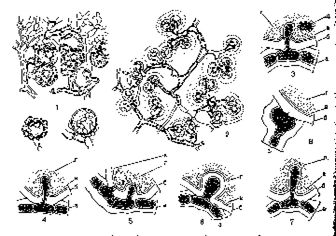
Более успешным оказалось изучение форм контакта между гифами гриба и клетками водорослей в слоевищах лишайников. Как показали исследования с применением электронной микроскопии, в слоевище лишайников можно встретить по крайней мере пять типов контакта между грибными гифами и водорослевыми клетками (рис. 289).
Чаще всего отдельная клетка водоросли и клетка грибной гифы находятся в непосредственном контакте друг с другом. В таком случае гриб образует специальные абсорбционные, всасывающие органы, которые проникают внутрь водорослевой клетки или плотно прижимаются к ее оболочке.
В настоящее время среди абсорбционных органов гриба в слоевище лишайников различают несколько типов: гаустории, импрессории и аппрессории.
Формы контакта между гифами гриба и клетками водорослей в слоевище лищайников.
Гаустории— это боковые выросты гиф гриба, которые прорывают оболочку клетки водоросли и проникают в ее протопласт (рис. 289, 2). Обычно в клетке водоросли развивается одни гаусторий, но иногда их может быть и два. В слоевище лишайника гаустории встречаются в большом количестве и существуют продолжительное время. Было замечено, что в оболочках молодых гаусториев нет отложений целлюлозы, которая могла бы затруднять обмен между клеткой водоросли и гифой гриба. Старые гаустории почти всегда одеты довольно толстым слоем целлюлозы. Различают интрацеллюлярные (внутриклеточные) и интрамембранные (внутриоболочковые) гаустории.
Интрацеллюлярные гаустории полностью прорывают оболочку клетки водоросли и проникают глубоко внутрь ее протопласта (рис.289, 3). Интрацеллюлярные гаустории образуются в случае резкого паразитизма гриба на водоросли. Это особенно характерно для лишайников с примитивным строением слоевища.
У более высокоорганизованных лишайников образуются только интрамембранные гаустории. Они прорывают оболочку клетки водоросли и достигают ее протопласта, но не углубляются в него, а остаются в оболочке водорослевой клетки (рис. 289, 5). Наибольшее количество интрамембранных гаусториев образуется в слоевище лишайников весной, в начале вегетационного периода. С наступлением осени они далеко отступают от протопласта водоросли.
Другой тип всасывающих органов гриба — импрессории— тоже боковые выросты грибных гиф, но, в отличие от гаусториев, они не прорывают оболочку клетки водоросли, а вдавливают ее внутрь (рис. 289, 6, 7). Импресcopии отмечены у очень многих лишайников, например у широко распространенной пельтигеры (Peltigera).
Интересно, что в слоевищах, произрастающих во влажных местообитаниях, импрессории почти не развиваются, у тех же видов в сухих местообитаниях они образуются в большом количестве. При длительной засухе число импрессориев также увеличивается. Предполагают, что в засушливые периоды и в сухих местообитаниях гриб, чтобы удовлетворить потребности в питании, увеличивает свою всасывающую поверхность за счет увеличения количества и размеров импрессориев.
В отличие от гаусториев и импрессориев, образованных боковыми отростками гифы, аппрессории образуются вершиной грибной гифы. Такая вершина гифы плотно прижимается снаружи к оболочке клетки водоросли, никогда не проникая ни в ее протопласт, ни в ее внутренний слой (рис. 289, 8).
Наличие в слоевищах многих лишайников абсорбционных органов гриба хорошо доказывает паразитическую сущность отношениймикобионта к фикобионту. Но во многих случаях у лишайникового гриба все же не удается обнаружить особых абсорбционных органов, чаще всего у лишайников, фикобионт которых имеет тонкие оболочки клеток. В таких случаях уже внешний контакт гифы гриба и клетки водоросли может обеспечить обмен веществами между ними. Так, например, обстоит дело у многих видов рода кладония. Фикобионтом кладонии является одноклеточная зеленая водоросль требуксия. У этих лишайников отдельные клетки водорослей окружены со всех сторон тонкими тонкостенными гифами, иногда поделенными на мелкие клеточки. Эти гифы, которые носят название обволакивающих или контактных, не проникают в протопласт клеток водоросли и не внедряются в их оболочку, а просто окружают клетки со всех сторон, так что каждая из них становится похожей на маленький шар, охваченный пальцами рук (рис. 289,7). Иногда гифы полностью оплетают водоросли в виде сплошного покрова и при этом, сливаясь своими стенками, даже образуют клеточную псевдопаренхимную ткань. На первый взгляд кажется, что водоросли не особенно страдают от плотного окружения гифами гриба: они долго сохраняют свою зеленую окраску и продолжают интенсивно делиться.
Но в более старых участках слоевища можно найти немало отмерших обесцвеченных клеток — гриб рано или поздно все-таки убивает водоросли. Такой же тип контакта между гифами гриба и клетками водорослей был найден у некоторых слизистых и базидиальных лишайников.