Реферат: Состав мембран
Основными компонентами мембран являются белки и липиды. На долю углеводов может приходиться около 10% массы мембран, при этом они всегда входят в состав гликолипидов или гликопротеи-нов. Соотношение между белками и липидами в мембранах значительно варьирует — от 20% белка в миелине до 80% в митохондриях. В табл. 1.3 и 1.4 обобщены данные по составу ряда мембран. Плотность мембран прямо пропорциональна содержанию в них белка. Судя по данным изопикнического центрифугирования, чем выше содержание белка в мембране, тем больше ее плотность.
Таблиця 1. Фосфолипидный состав субклеточных мембран печени крысы "
| Доля от суммарного количества фосфолипидов, It | ||||||
| митохондрии | микро-сомы |
ли10- сомы | плазматическая мембрана | ядерная мембрана | мембраны аппарата Гольджи | |
| Кардиолипин | 18 | 1 | 1 | 1 | 4 | 1 |
| Фосфатидилэтаноламин | 35 | 22 | 14 | 23 | 13 | 20 |
| Фосфатидилхолин | 40 | 58 | 40 | 39 | 55 | 50 |
| Фосфатидилииозитол | 5 | 10 | 5 | 8 | 10 | 12 |
| Фосфатидилсерин | 1 | 2 | 2 | 9 | 3 | 6 |
| Фосфатидная кислота | — | 1 | 1 | 1 | <1 | |
| Лиэофосфоглицериды 2 ' | 1 4 | 11 | 7 | 2 | 3 | 3 |
| Сфингомиелин | 1 | 1 | 20 | 16 | 3 | 8 |
| Фосфолипиды | 0,175 | 0,374 | 0,156 | 0,672 | 0,500 | 0,825 |
| Холестерол | 0,003 | 0,014 | 0,038 | 0,128 | 0,038 | 0,078 |
Таблица 2 Белковый и липидиый состав некоторых мембран животных и бактериальных клеток. Л/Б — отношение липка/белок"

Белковый состав мембраны зависит в какой-то степени от метода ее выделения. Некоторые белки непрочно связаны с мембраной и легко удаляются при промывании ее растворами с высокой или, напротив, с низкой ионной силой, щелочными растворами или растворами, содержащими хелатирующие агенты типа ЭДТА. Бывают и случаи, когда трудно сказать, является ли белок мембранным или цитоплазматическим, случайно связавшимся с мембраной в ходе ее выделения.
1. МЕМБРАННЫЕ ЛИПИДЫ
Наиболее поражает в мембранных липидах их огромное разнообразие. Причины этого пока не ясны, хотя становится все более очевидно, что, по-видимому, связано это с тем разнообразием функций, которые липиды выполняют в мембранах. Но, конечно, главная функция мембранных липидов состоит в том, что они формируют бислойный матрикс, с которым взаимодействуют белки. Основные классы липидов представлены на рис. 1.8; их краткое описание дано ниже.
Глицерофосфолипиды
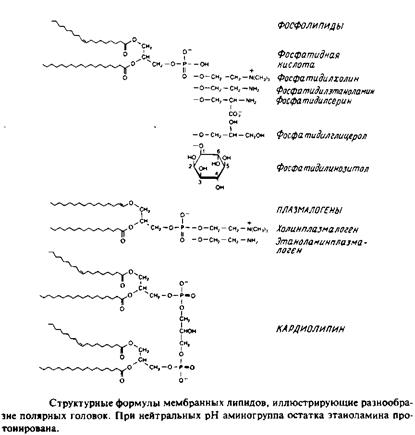
Это наиболее распространенные липиды. Одна из гидроксильных групп глицерола связана с полярной группировкой, содержащей фосфат, а две другие — с гидрофобными остатками. Номенклатура гли-церидов основана на системе стереоспецифической нумерации. Если глицерол изобразить в проекции Фишера, так что центральная группа будет расположена слева, то атомы углерода будут нумероваться так, как показано на рис. В этом случае в название глицерофосфолипида вводят приставку snдля обозначения положения заместителя. В литературе встречается несколько систем стереохимических обозначений: sn, D/L и R/S. На рис. представлена стереохимия атома С-2 в этих трех системах. Природные фосфолипиды.
У большинства фосфоглицеридов фосфатная группа находится в ял-3-положении глицерола; она обычно связана с какой-либо из групп, представленных на рис.
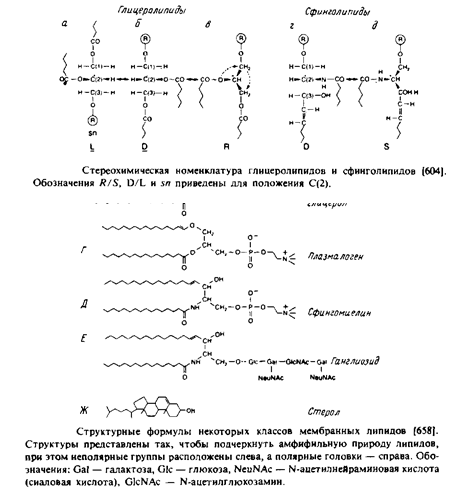
Длинные углеводородные цепи, находящиеся в положениях sn-\ и sn-2, могут присоединяться за счет сложноэфирной и простой эфирной связей. Эти цепи значительно различаются по длине, раз-ветвленности и степени ненасыщенности.
1. 1,2-диацилфосфоглицериды или фосфолипиды. Эти липиды, являющиеся сложными эфирами жирных кислот и глицерола, широко представлены во многих мембранах эукариотических и прокарио-тических клеток, за исключением архебактерий. Фосфатидил-холин является основным компонентом мембран животных клеток, а фосфатидилэтаноламин — это нередко основной липид бактериальных мембран. В табл. представлен ряд жирных кислот, наиболее часто встречающихся в составе фосфолипидов, а в табл. приведен жирнокислотный состав клеточных мембран печени крысы.

Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. Степень ненасыщенности может быть разной, но чаще всего встречаются ненасыщенные кислоты 18:1, 18:2, 18:3 и 20:4. Здесь первое число обозначает длину ацильной цепи, а второе указывает на число содержащихся в ней двойных связей. Почти все природные кислоты характеризуются г/ис-конфигурацией двойных связей. Цепь в такой конфигурации имеет излом, что нарушает упаковку липидных молекул в бислое. В составе молекул многих фосфолипидов имеется одна насыщенная и одна ненасыщенная цепи. В случае животных клеток ненасыщенные цепи обычно находятся в 5и-2-положении глицерола. Такое же распределение цепей характерно и для фосфолипидов клеток Е. coli. Двойные связи в полиненасыщенных цепях обычно являются несопряженными. В фосфолипидах некоторых бактериальных мембран обнаружены разветвленные цепи, а также цепи, содержащие циклы, и гидроксильные группы в /3-положении. На рис. 1.11 показана структура некоторых из этих кислот.
2.У архебактерий глицерофосфолипиды имеют обращенную сте-реохимическую конфигурацию, при которой фосфорильные группы находятся в sn-\-положении глицерола. У многих бактерий этого вида гидрофобные компоненты представляют собой не сложные эфиры жирных кислот, а простые изопранильные эфиры глицерола.
Кардиолипины или дифосфатидилглицеролы. По сути это димерные формы фосфолипидов. Они содержатся в боль шом количестве во внутренней мембране митохондрий, в мембране хлоропластов и в некоторых бактериальных мембранах, но редко встречаются в других мембранах.

4. Плазмалогены. Это фосфоглицеролипиды, у которых одна из углеводородных цепей представляет собой простой виниловый эфир. Этаноламиновые плазмалогены широко представлены в миелине и в саркоплазматическом ретикулуме сердца.
Фосфосфинголипиды
Эти липиды имеют такие же полярные головки, как и глицерофосфолипиды, но их гидрофобная часть представлена церамидом. В плазматических мембранах животных клеток широко распространен сфингомиелин. Основными жирнокислотными компонентами в миелине являются кислоты 24:1 и 24:0. В мембранах растительных и бактериальных клеток фосфосфинголипиды встречаются редко. Кроме сфингомиелина известны и другие фосфосфинголипиды, например церамид-1-фосфорилэтаноламин, церамид-1-фосфорилинози-тол и церамид-1-фосфорилглицерол.
Гликоглицеролипиды
Это полярные липиды, у которых в м-З-положении глицерола находится углевод, присоединенный с помощью гликозидной связи, например галактоза. Гликоглицеролипиды широко представлены в мембранах хлоропластов, они обнаружены также в заметных количествах в сине-зеленых водорослях и бактериях. Моногалактозилдиа-цилглицерол был назван «наиболее распространенным в природе полярным липидом», поскольку на его долю приходится половина всех липидов тилакоидной мембраны хлоропластов. Для мембран грамположительных бактерий характерны гликоглице-
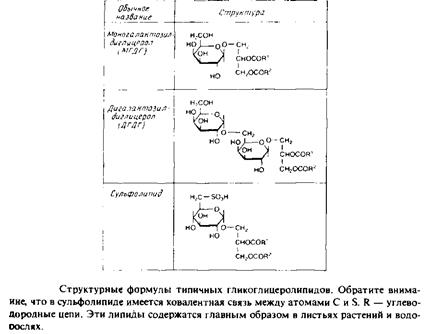
ролипиды с большим разнообразием Сахаров. Архебактерии также содержат такие липиды, но, как и в случае глицерофосфолипидов, их стереохимическая конфигурация является обращенной, с локализацией гликозидной связи в sn-l-положении глицерола. В мембранах животных клеток гликоглицеролипиды встречаются редко.

Гликосфинголипиды
классифицируют в соответствии с размером углеводной части, которая может быть представлена всего лишь одним моносахаридным остатком, с одной стороны, и очень сложным углеводным полимером — с другой. Моногликозилцерамиды обычно называют цереброзидами. Ганглиозиды представляют собой класс анионных гликосфинголипидов, которые содержат один или несколько остатков сиаловой кислоты, связанных с сахарными остатками церамидолигосахарида. Глобозидами называют нейтральные гликосфинголипиды, которые не содержат остатков отрицательно заряженной сиаловой кислоты.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--