Реферат: Структура и свойства мембранных липидов
Параметры можно определить по данным дифракции рентгеновских лучей. Как правило, те параметры, которые зависят главным образом от длины ацильных цепей, почти не меняются при увеличении содержания воды в системе. Гидратация липидов происходит в результате связывания воды с полярными головками. Процесс гидратации активно изучали методами 'Н- и 2 Н-ЯМР. Результаты, полученные при изучении фос-фатидилглицерола, фосфатидилхолина и фосфатидилэтаноламина методом 2 Н-ЯМР, говорят о наличии гидратной оболочки из 11 — 16 молекул воды на одну молекулу липида. Эти молекулы быстро обмениваются с молекулами воды, находящимися в основном объеме. Согласно другим измерениям, полярная головка фосфатидилэтаноламина связывает меньше воды, чем полярная головка фосфатидилхолина. Было высказано предположение, что в случае ненасыщенных фосфатидилэтаноламинов слабая гидратация благоприятствует образованию неламеллярной гексагональной фазы Ни.
Многие липиды набухают в воде. Липиды, которые не несут заряда или являются в целом электрически нейтральными, не набухают совсем или набухают лишь в ограниченной степени до предельной толщины водной прослойки между ламеллами. При избытке воды сосуществуют две фазы — мультиламеллярная липидная фаза и вода, находящаяся в основном объеме. Заряженные липиды склонны к неограниченному набуханию и могут включать воду между ламеллами вплоть до пороговой точки, когда образуются две фазы — полностью гидратированные моноламелярные везикулы, находящиеся в равновесии с водой в основном объеме. Степень набухания и относительная стабильность мульти- и моноламеллярных структур определяются электростатическими взаимодействиями. При низкой ионной силе происходит дестабилизация мультиламеллярных структур. Неограниченное набухание может происходить и в том случае, когда в смеси липидов содержится всего несколько процентов заряженных липидов.
Поляризация молекул воды вблизи полярных липидных головок приводит к сильному отталкиванию при сближении двух бислоев. Эта «гидратационная сила» удерживает гидратированные бислои на расстоянии не менее 30 А друг от друга. Именно она создает основной энергетический барьер, который следует преодолеть, пытаясь осуществить слияние мембран. Возможно, фосфатидилэтаноламиновые везикулы склонны к агрегации именно потому, что степень гидратации их полярных головок относительно низка.
Исследования, проведенные методом ЭПР с помощью спиновых меток, способных реагировать на полярность своего окружения на различной глубине от поверхности бислоя, показывают, что вода частично проникает в углеводородную область бислоя, находящуюся в жидкокристаллическом состоянии. По данным нейтронного рассеяния, когда бислой находится в состоянии геля, вода не проникает глубже глицеролового остова липидных молекул.
2.2 ФАЗОВЫЕ ДИАГРАММЫ ОДНОКОМПОНЕНТНЫХ ВОДНО-ЛИПИДНЫХ СИСТЕМ
На рис. 2.5 представлена фазовая диаграмма в координатах температура—концентрация для двух распространенных гликолипидов, выделенных из растений. Дигалактозилдиацилгли-церол образует стабильную ламеллярную фазу. При низком содержании воды имеется только гидратированная фаза, а при содержании воды 20% сосуществуют две фазы — мультиламеллярная гидратированная липидная фаза и вода основного объема. К неограниченному набуханию ДГДГ не способен.
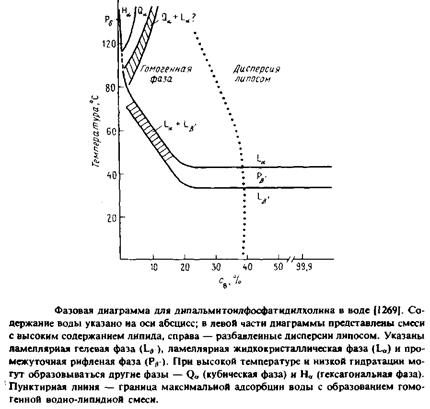
На приведена фазовая диаграмма для дипальмитоилфос-фатидилхолина. Этот липид существует в ламеллярной форме при самых разных условиях. При добавлении воды он «набухает» до тех пор, пока между слоями не скопится максимально возможное количество воды. В этой точке образуется двухфазная система, представляющая собой взвесь мультиламеллярных липо-сом. Обратите внимание, что между фазой геля и жидкокристаллической ламеллярной фазой имеется еще одна фаза. Это так называемая «рифленая» фаза. В этой фазе поверхность бислоя на электронных микрофотографиях имеет волнообразный вид. Температурный фазовый переход Р0 , -» La называется главным, а переход ц, -» Р^, — предпереходом.
Липиды, имеющие объемные полярные головки, обычно претерпевают предпереход, свидетельствующий о наличии фазы, промужеточной между гелевой и жидкокристаллической. В Ра ,-фазе дипальмитоилфосфатидилхо-лина ацильные цепи, вероятно, наклонены, хотя спектры комбинационного рассеяния говорят о том, что они по-прежнему, как и в фазе геля, находятся преимущественно в полностью-трянс-конфигурации.
2.3 ДВА МЕТОДА ИЗУЧЕНИЯ ЛИПИДНОГО ПОЛИМОРФИЗМА
Кроме метода дифракции рентгеновских лучей для характеристики свойств липидных фаз использовали и другие методы. Один из них, оказавшийся особенно полезным, — это электронная микроскопия замороженных сколов. Второй метод, 31 Р-ЯМР, был введен в исследовательскую практику несколько позже и нашел применение для обнаружения небислойных структур, которые, как полагают, играют особую роль в биологических мембранах.
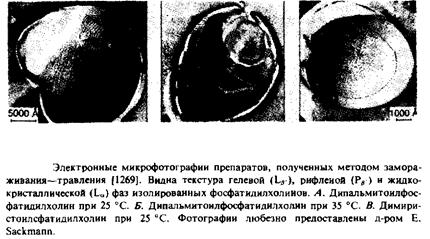
1. Электронная микроскопия липидных фаз с применением метода замораживания—травления. Этот метод оказался очень полезным при изучении структурной организации различных липидных фаз. Примеры получаемых при этом электронных микрофотографий приведены на рис. 2.7. Жидкокристаллическая фаза на этих микрофотографиях всегда выглядит как гладкая поверхность, а Р^.-фаза — как рифленая. Геле-вая фаза выглядит как гладкая поверхность, но при определенных условиях приготовления препарата имеет спиральную текстуру из-за возникновения случайных дефектов в плотной упаковке. Гексагональная фаза, образуемая такими лип идами, как ненасыщенный фосфатидилхолин, выяляется на микрофотографиях после замораживания—травления как стопка цилиндров. При приготовлении образцов липиды уравновешивают в условиях, подходящих для формирования нужных структур, а затем быстро замораживают, чтобы организация этих структур не успела измениться.
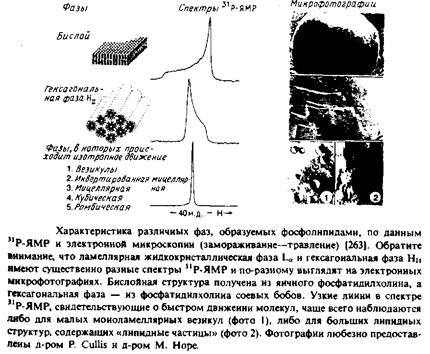

Электронно-микроскопические методы использовались также для изучения «липидных частиц». Эти частицы нередко наблюдаются в бинарных смесях, если один из липидов склонен образовывать фазу Ни, а другой — бислойные структуры. Липидные частицы образуются в чисто липидных образцах и потому отличаются от «белковых частиц», которые наблюдаются в биологических мембранах и в реконструированных белково-липид-ных системах. Было высказано предположение, что липидные частицы представляют собой вывернутые мицеллы, расположенные внутри бислоя, и что они играют определенную роль в биологических процессах, облегчая слияние мембран или стабилизируя их на сильно искривленных участках, как, например, в мем^ бране тилакоидов. Однако четких доказательств биологической роли липидных частиц пока не получено.
2. Исследование бислоев методом п Р-ЯМР. Этот метод также используется для структурной характеристики гидратирован-ных липидов. Например, фосфолипиды в гексагональной фазе Ни дают спектры 31 Р-ЯМР, резко отличающиеся от спектров фосфолипидов в ламеллярной фазе. С помощью этого метода исследовался переход ламеллярной фазы в гексагональную в липидах и липидных смесях. Недостатком метода является некоторая неопределенность при интерпретации спектров в случае изотропного усреднения за счет относительно быстрых движений. Полагают, что такой спектр отвечает структурам типа «липидных частиц», однако это объяснение не является единственно возможным. Как было показано, метод 31 Р-ЯМР позволяет надежно установить наличие гексагональной Ни-фазы в чисто липидных дисперсиях, но в случае биологических мембран к подобным выводам следует относиться с осторожностью, особенно если они не подтверждены другими методами.
Метод 31 Р-ЯМР оказался весьма полезным при определении ориентации и динамического поведения полярных головок фосфолипидов, а также структурных возмущений, вносимых в бислой мембранными белками.
2.4. ОРИЕНТАЦИЯ ПОЛЯРНЫХ ГОЛОВОК ЛИПИДОВ В БИСЛОЕ
Данные ряда методов свидетельствуют о том, что в ламелляр-ных водно-фосфолипидных дисперсиях, как и в липидных кристаллах, полярные головки липидов в целом ориентированы параллельно плоскости бислоя. В случае фосфатидилхолинов такая ориентация присуща как гелевой, так и жидкокристаллической фазам, судя по результатам исследований методами дифракции нейтронов и рентгеновских лучей. Об этом же свидетельствуют и данные, полученные методом 2 Н-ЯМР, хотя при этом не исключаются и другие интерпретации. Имеющиеся данные указывают на то, что пространственное расположение полярных головок фосфа-тидилглицерина, сфингомиелина и фосфатидилсерина сходно. Исследования методом 2 Н-ЯМР интактных фибробластов мыши и выделенных из них мембран также показали, что полярные головки как фосфатидилхолина, так и фосфатидилэтаноламина ориентированы параллельно поверхности мембран. Однако по данным дифракции нейтронов у фосфатидилглицерола, выделенного из Е. coli, полярная головка ориентирована примерно под углом 30° к поверхности мембраны, что облегчает связывание отрицательно заряженных фосфатных групп с катионами.
На ориентацию и динамику полярных головок липидных молекул может влиять образование межмолекулярных водородных связей на поверхности мембраны. Донорами и акцепторами при Образовании этих связей могут служить такие липиды, как фосфатидилсерин, фосфатидилэтаноламин и различные гликолипиды. Исследования, проведенные на модельных мембранных системах, показывают, что водородные связи между полярными головками сохраняются даже в условиях гидратации мембранной поверхности, однако пока неизвестно, как образование этих связей может сказаться на структуре биологических мембран.
2.5. КОНФИГУРАЦИЯ И УПАКОВКА АЦИЛЬНЫХ ЦЕПЕЙ В БИСЛОЕ
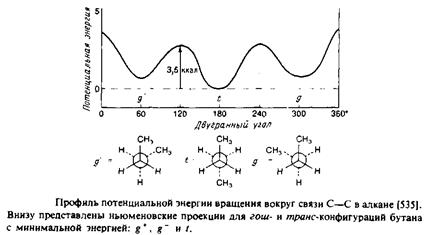
Рассмотрим сначала насыщенные углеводородные цепи. В них возможно свободное вращение вокруг каждой С—С-связи, характеризующееся энергетическим минимумом, особенно четким в случае ньюменовской проекции. Наиболее стабильна транс-конфигурация, при этом высота энергетического барьера для перехода через заслоненную конфигурацию в гош-форму составляет по оценкам 3,5 ккал/моль. В полностью m/wwc-конфигурации цепь максимально вытянута и не меняет своего направления, тогда как в гош-конформации ее направление меняется. Последовательность гош-транс-гош для трех смежных С—С-связей приводит к появлению в цепи излома, в результате чего участки цепи выше и ниже места излома оказываются значительно смещенными друг относительно друга. /ош-конфигурация в зависимости от направления вращения при переходе от Qк С4 обозначается как g+ или g". Кинки типа g+ tg~ или g~ tg+ приводят к минимальному сдвигу цепи. Почти все двойные связи в мембранных липидах находятся в J/ис-конфигурации. Как и в случае гош-конфигурации, это приводит к изменению общего направления цепи. Наличие в углеводородных цепях кинков, двойных ^ыс-связей,
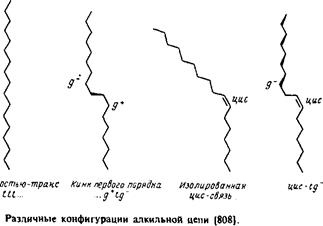
циклопропановых групп и других особенностей приводит к увеличению площади поперечного сечения цепи; это может иметь важные последствия для упаковки липидов в бислое. При этом стерические требования к упаковке углеводородных цепей и полярных головок такие же, как и в липидных крастал-лах. Эти принципы будут обсуждаться в разд. 2.3 при анализе формы мицелл.
Многие методы, включая дифракцию рентгеновских лучей и нейтронов, спектроскопию КР и ИК-спектрометрию, указывают, что в фазе геля насыщенные углеводородные цепи фосфолипидов находятся преимущественно в полностью-гпрднс-конфигурации. Минимальная площадь поперечного сечения молекулы диа-цильного фосфолипида равна около 38 A2 . Примерно такую площадь занимает полярная головка фосфатидилэтанола-мина, поэтому насыщенные фосфатидилэтаноламины в гелевой фазе упаковываются так, что ацильные цепи располагаются перпендикулярно плоскости бислоя, как и в липидных кристаллах. В случае же кристаллов фосфатидилхолина минимальная площадь, приходящаяся на одну полярную головку, составляет примерно 50 А2 . Поэтому дипальмитоилфосфатидилхолин в гелевой фазе не может упаковываться так, как фосфатидилэтаноламин. В этом случае ацильные цепи дипальмитоилфосфатидилхолина отклоняются на 30° от нормали к бислою, благодаря чему их поперечное сечение увеличивается и достигается соответствие размеру полярной головки. При этом углеводородные цепи сохраняют полнос-тью-транс-конфигурацию. В жидкокристаллической фазе появление в цепи гош-конформеров увеличивает эффективное поперечное сече-
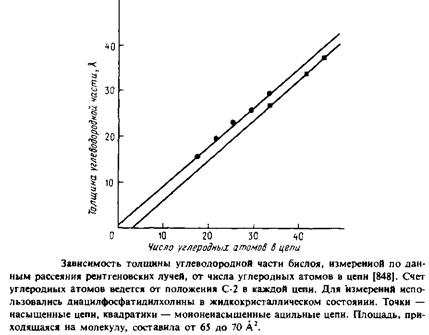
ние цепей по меньшей мере до 50 А2 в расчете на молекулу диациль-ного фосфолипида, а в водных дисперсиях эффективная площадь, приходящаяся на молекулу фосфолипида, составляет обычно 60 — 70 А2 . Следовательно, в жидкокристаллической фазе углеводородные цепи не наклонены к плоскости бислоя, поскольку в этих условиях.полярные головки липидных молекул достаточно удалены друг от друга, и чтобы заполнить пространство между соседними головками и перекинуть между ними мостики, требуются вода и другие полярные молекулы. Судя по данным 2 Н-ЯМР, толщина углеводородной области дипальмитоилфосфатидилхолинового бислоя в жидкокристаллическом состоянии составляет 35, а не 45 А, как следовало ожидать, если бы цепи находились в полностью-/я/*7«с-конфиругации и были ориентированы вдоль нормали к би-слою. Толщина бислоя уменьшается за счет наличия к цепях гош-конформеров, приводящих к разупорядоченности цепей, причем сами цепи в целом растянуты и расположены перпендикулярно поверхности бислоя, а не скручены в спираль. На рис. 2.12 приведена линейная зависимость толщины жидкокристаллического бислоя в диацилфосфатидилхолиновых везикулах от длины ацильных цепей, построенная по данным рассеяния рентгеновских лучей.
МЕТОДЫ ИССЛЕДОВАНИЯ ГИДРОФОБНОЙ ОБЛАСТИ БИСЛОЯ
Для получения детальной картины строения внутренней области бислоя особенно полезными оказались два метода: Н-ЯМР и ИК-и КР-спектроскопия. Применение *Н- 13 С-ЯМР для исследования мембран и водно-липидных систем затруднено, поскольку для получения пригодных для исследования малых везикул или мембранных фрагментов необходима предварительная фрагментация мембранного препарата с помощью ультразвука. Впрочем, если вращать образец под магическим углом, то необходимость в такой обработке отпадает, и это открывает новые возможности для более широкого применения последних методологических достижений ЯМР при изучении модельных и биологических мембран.
Применение методов 2 Н-ЯМР и колебательной спектроскопии не сопровождается возмущениями бислоя, поскольку в этих методах не используются зонды, которые могли бы исказить структуру окружающих их липидов. Ниже приведены краткое описание этих методов и сводка результатов, полученных при изучении конфигурации ацильных цепей.
Спектроскопия комбинационного рассеяния
Лазерная спектроскопия комбинационного рассеяния оказалась весьма полезной для изучения физического состояния модельных и биологических мембран. Этот метод основан на измерении разности энергии падающего света и света, рассеянного за счет колебательных движений. Тип колебаний и их интенсивность очень сильно зависят от физического состояния липидов, поэтому спектры фосфолипидов в фазе геля и жидкокристаллическом состоянии существенно различаются. Особенно полезны для регистрации этих различий валентные колебания связи С—С. Например, появление гош-конформеров при плавлении углеводородных цепей приводит к увеличению интенсивности полосы при 1080 см-1 . Этот метод показывает, что некоторая доля гош-конформеров сохраняется и в состоянии геля до тех пор, пока температура не снизится до очень низких значений, около - 200° С. Заметим, что колебания часто охватывают всю молекулу, так что количественная интерпретация спектров с точки зрения анализа влияния отдельных коиформаций является непростой задачей.