Реферат: Зависимость интенсивности дыхания растительных продуктов от температуры
К группе Сu-протеидов относятся различные полифеноло-ксидазы и аскорбиноксидаза. В основе их действия лежит обратимое окисление одновалентной меди в двухвалентную.
Полифенолоксидазы представляют собой ферменты, окисляющие в присутствии молекулярного кислорода различные фенолы и их производные с образованием соответствующих хинонов. Полифенолоксидазы катализируют прямое окисление субстратов атмосферным кислородом и участвуют в различных окислительных процессах, в том числе в окислении аминокислот, полипептидов, белков и аскорбиновой кислоты. Активность полифенолоксидаз возрастает при механическом повреждении плодов, воздействии низких температур, кристаллизации воды при замораживании и т. д.
Потемнение мякоти многих плодов (яблок, груш, абрикосов и др.), а также клубней картофеля, наблюдаемое при разрезании, очистке и механическом повреждении, происходит вследствие возрастания активности полифенолоксидаз и нарушения координации реакций окисления и восстановления, в результате чего в ткани накапливаются стойкоокрашенные продукты.
Аскорбиноксидаза обладает строгой специфичностью по отношению к субстрату и окисляет l-аскорбиновую кислоту, превращая ее в дегидроформу. Являясь переносчиком водорода, аскорбиновая кислота тесно связана со всей системой ферментов, участвующих в дыхательных процессах.
Полифенолоксидазы и аскорбиноксидаза принимают участие в окислительных процессах на заключительном этапе дыхания, заменяя цитохромы.
На рис. 2 показаны возможные альтернативные пути процесса окисления, которые в. растительных клетках происходят различным образом в зависимости от видовых особенностей ткани, физиологического состояния данного органа, условий внешней среды и других факторов.
Различная активность оксидаз в условиях измененной газовой среды представлена на рис. 3.
Наибольшее сродство к кислороду имеет цитохромоксидаза, ее действие проявляется при содержании кислорода в атмосфере всего 1 %. Глубинные слои паренхимной ткани плодов и овощей, для которых характерно пониженное содержание кислорода, более богаты цитохромоксидазой, нежели поверхностные.
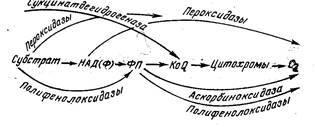
Рис. 2. Альтернативные пути окисления
Ферменты, участвующие в альтернативных процессах окисления, локализованы в различных структурных элементах клетки. Пероксидазы, полифенолоксидазы, аскорбиноксидаза сосредоточены главным образом в цитоплазме; вся система ферментов, обеспечивающая окислительное фосфорилирование, обнаружена в ядрах и хлоропластах. Благодаря такой делокализации ферментов дыхательной цепи практически все структурные составляющие клетки могут быть источниками того или иного количества энергии и использоваться ею в зависимости от условий внешней среды.
О характере и интенсивности дыхания судят по количеству выделяемого клетками углекислого газа либо по величине дыхательного коэффициента ДК:
ДК=Vсо,/Vо„ где Vco , Vo - объемы выделившегося углекислого газа и поглощенного кислорода.
Величина дыхательного коэффициента определяется прежде всего химической природой субстрата. При окислении углеводов ДК=1 в соответствии с суммарным уравнением окисления глюкозы
С Н О + 60 -> 6СО + 6Н О + Q.
Число выделившихся грамм-молекул углекислого газа равно числу поглощенных грамм-молекул кислорода, а согласно закону Авогадро грамм-молекулы всех газов занимают одинаковый объем.
При дыхании число выделившихся молекул углекислого газа равно числу атомов углерода в молекуле субстрата. Поэтому число использованных молекул кислорода на атом углерода в субстрате возрастает с увеличением числа Н-атомов и снижается с увеличением содержания O-атомов в молекуле субстрата.
Если субстратами дыхания являются высшие карбоновые кислоты или аминокислоты, относительно богатые водородом и бедные кислородом, то ДK<1. Для жиров ДК~0,7, для белков ДК~0,8. В качестве примера приведем окисление стеариновой кислоты:
С Н О + 26O -»18СО + 18Н О; ДК = 18/26 = 0,69.
Если дыхание происходит с использованием низших ди- и трикарбоновых кислот (яблочной, лимонной, винной и щавелевой), богатых кислородом, то ДК>1.
Так, при окислении яблочной кислоты
С Н О + ЗО -» 4СО + ЗН О; ДК. = 4/3 = 1,33.
Интенсивность дыхания и соответственно величина дыхательного коэффициента молодых развивающихся тканей, например меристематических, тканей прорастающих семян выше, чем зрелых плодов. У тканей одного и того же плода или овоща интенсивность дыхания периферических тканей, как правило, выше.
Ниже приведена интенсивность дыхания [в мг СO2/(г*сут)] различных тканей при температуре 15—20 °С.
|
12,4 69,3 10,6 2,45 11,3 13,8 3,8 |
Лимон
целые плоды