Статья: Морфологическая изменчивость у колониального гидроида dynamena pumila и классификация обнаруженных морфотипов книдарии
Разработан метод анализа морфологической изменчивости популяции колониального гидроида Dynamena pumila на основе учета отклонений от нормы формы перисарка стереотипных междоузлии побегов, называемых морфологическими аномалиями или морфотипами. По результатам обработки более 200 000 междоузлии составлена классификация морфологических аномалий, включающая 7 разделов и 47 разновидностей. В статье представлены описания и рисунки всех морфотипов. Материал собран на Белом и Баренцевом морях в 1991—1993 гг.. а также 100 лет тому назад (коллекции Зоологического института РАН). Некоторые морфотипы выглядят как полноценные резервные фенотипы вида. Для большинства морфотипов приведены объяснения возможных причин их образования.
Из всех гидроидов пока что только для одного вида описана изменчивость строения стереотипных частей колонии — междоузлии побегов (Марфенин, 1975, 1988), Этот вид—Dynarnena pumiia (L.) из высокоорганизованного семейства Sertulariidae отряда Thecaphora — обладает в норме двурядным супротивным расположением гидрантов на стволе побега (рис. 1). В процессе роста побега очередная пара гидрантов формируется на его верхушке. Хитиновый перисарк, выделяемый здесь, принимает во время морфогенеза верхушки соответствующую форму, имеющую видоспецифический характер. Морфогенез побега протекает циклично, так что к концу образования очередной пары гидрантов воссоздаются исходные условия для формирования следующей пары (рис. 2). Каждый стереотипный участок побега, включающий пару гидрантов и отрезок ствола, ясно отграничен от следующего подобием перетяжки, что позволяет выделять в побеге соответствующие междоузлия.
Таким образом, типичный для Dynamena pumiia морфогенез повторяется в каждой колонии множество раз, что позволяет изучать фенотипическую изменчивость даже в пределах отдельно взятого генетически однородного организма, каковым на самом деле и является колония у гидроидов (Марфенин, 1993). Подобную задачу можно ставить и на других так называемых модульных организмах, вроде растений и микромицетов, или же на совокупности клеток одного организма, однако до сих пор такие исследования единичны.
Возможно, это обстоятельство объясняется излишней вариабельностью строения модулей (листьев, побегов),т.е. недостаточной стабильностью их канонической формы, что затрудняет выделение четких критериев для исследования внутривидовой изменчивости. Вторая возможная причина связана с недостаточностью проявления вариабельности строения модулей у многих видов. Например, у гидроидов из семейства Campanulariidae нам не приходилось встречать обращающих на себя внимание морфологических отклонений от нормы в строении частей колонии.
Dynamena pumila в этом отношении значительно отличается от других гидроидов, так как ненормальные по своему внешнему виду междоузлия встречаются в ее колониях довольно часто. По существу у D. pumila мы встречаем разнообразие морфотипов. представленное как доминирующей формой междоузлия, принятой за норму, свойственную данному виду, так и значительно реже встречающимися другими формами. В большинстве случаев они выглядят уродливо и поэтому первоначально были названы аномалиями формы побега (Марфенин, 1975). Однако среди аномалий встречаются морфологически и функционально вполне полноценные формы, которые могут последовательно и относительно устойчиво воспроизводиться в побеге. И те, и другие отклонения от канонической формы обладают постоянными индивидуальными чертами строения, отличными друг от друга, т. е. присущи виду, поэтому правильнее их в общем случае называть морфотипами, хотя для многих явно уродливых образований мы оставляем название морфологические аномалии.
Полноценному изучению феномена внутриколониальной морфологической изменчивости Dynamena pumila должны предшествовать описание и систематизация морфотипов, чему и посвящена настоящая статья. Если первоначально были описаны 27 морфотипов неканонического строения междоузлии побега D. pumila (Марфенин, 1975), то после специального исследования тысяч побегов нами выделено множество аномалий, которые сведены к 47 морфотипам.
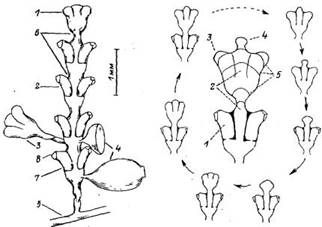
Рис. 1. Схема строения нормального побега Dynamena pumila:
1 — верхушка роста ствола побега; 2 — ствол побега; 3 — боковая ветвь побега (7 порядка); 4 — гонотенн на разных стадиях развития; 5 — столон; б — перетяжки побега (границы неждоузлий; 7— гидротека (нижняя часть гидротеки — диафрагма); 8—устье гидротеки
Рис. 2. Морфогенетический цикл образования междоузлия у побега Dynaтепа pumila.
I — нормальная гидротека сформировавшегося гидранта; 2 — ствол побега; 3 — формирующийся гидрант; 4 — верхушка роста на ранней стадии морфогенеза; 5—последовательные характерные стадии развития верхушки
Материал и методика
К настоящему времени удалось обстоятельно изучить две популяции Dynamena pumila: одна из Ругозерской и Кислой губ Кандалакшского залива Белого моря, а другая из губы Дальне-зеленецкой Баренцева моря. Пробы были собраны летом 1991 и 1992 гг. на Белом море нами, а на Баренцевом море по нашей просьбе сотрудницей ММБИ К. Н. Пантелеевой (за что приносим ей искреннюю благодарность).
Дополнительно был использован музейный материал из коллекции Зоологического института РАН, собранный в Белом море (о-в Сидоров) в 1895 г., а в Баренцевом море в 1893 г. (Екатерининская гавань) и в 1899 г. (о-в Седоватый). Сравнительный анализ изменения встречаемости морфотипов D. pumila за 100 лет будет рассмотрен в отдельной статье. В данном исследовании мы хотели составить как можно более полное представление о спектре присущих виду морфотипов по всему имеющемуся у нас материалу.
Для исследования отбирали малообросшие колонии с субстратом. Побеги отрезали у основания и фиксировали, или же фиксировали колонии вместе с субстратом. Следует иметь в виду, что в колониях могут быть уродливые крошечные побеги, которые при отборе более крупных остаются незамеченными и неучтенными. При обработке материала побеги просматривали под бинокуляром. В журнале регистрации записывали: 1) порядковый номер; 2) число междоузлии в стволовой части побега; 3) число междоузлии в каждой боковой ветви побега (сначала ветви 1 порядка, затем 11 порядка и т. д.); 4) суммарное число междоузлии и побеге; 5) номера морфотипов по описанной ранее классификации (Марфенин, 1975) или же номер рисунка, не попадающего эод прежнюю классификацию морфотипа; б) месторасположение лорфотипа (число междоузлии до аномалии), считая от верхушки побега. Все побеги с отклонениями от канонического морфотипа сохраняли в коллекции.
Правильнее всего оценивать выборки по суммарному числу обследованных междоузлии и лишь затем — по числу побегов. Зсего было исследовано более 200 тыс. междоузлии беломорской юпуляции и более 110 тыс. междоузлии баренцевоморской популяции D. pumila. Соответственно, более 8000 и 5000 побегов.
В конечном результате по каждой выборке можно было указать совокупную долю (%) всех неканонических морфотипов и встречаемость (%) каждого из них. В данной статье мы будем считать, ito морфотип встречается: а) очень редко, если доля его находок во всей совокупной выборке составляла менее 0.002%, 5) редко: от 0.01 до 0.002%, в) периодически: 0.05— 3.011%; г) часто: 0.1—0.051%; д) очень часто:более 0.1%
Регистрация расположения морфотипов в колонии не дала каких-либо интересных результатов. Аномалии распределялись в пределах побега достаточно равномерно.
Систематизация морфотипов представляет собой самостоятельную задачу, которую мы обсудим ниже, после основного раздела, содержащего их описания. Мы остановились на самом простом и достаточно формальном подходе: морфотипы условно разделены по группам в соответствии с их местоположением в колонии. Всего получилось 7 групп.
Цифровые обозначения присваиваются морфотипам по следующему порядку. Первая (римская) цифра обозначает номер группы. Через тире указывается арабскими цифрами присваиваемый порядковый номер морфотипа. Близкие по своему строению аномалии получают номера в пределах одного десятка. Пропущенные в пределах десятка номера резервируются для будущих находок и одновременно маркируют границы подгрупп. В том случае, когда аномалия была описана (Марфенин, 1975), в скобках приводится ее прежний номер. Если таковой отсутствует, морфотип описан впервые.
Описание различных отклонений от нормального морфотипа dynamena puмila
Группа I: Отклонения в морфогенезе ствола побега без остановки его дальнейшего роста (рис. 3)
Аномалии выражаются в необычном взаимном расположении междоузлии, гидрантов, а также необычной форме междоузлии. Форма гидротек остается нормальной.
I—1 (1), Изгиб ствола без деформации междоузлия может происходить в самые различные стороны относительно продольной оси побега. Наибольший угол отклонения равен 90°. Местом изгиба является исключительно перетяжка. Возникает после приостановки роста побега в том случае, если при начале очередного периода роста верхушка ценосарка растворяет хитиновую теку не апикально, а сбоку. Кроме того, аномалия возникает в том случае, если растущий побег встречает на своем пути преграду. Встречается периодически.
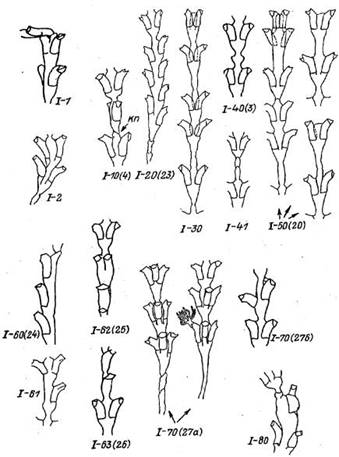
Рис. 3. Отклонения в морфогенезе ствола побега Dynamena pumUa без остановки его дальнейшего роста (группа I)
кп— косая перетяжка
1—2. Изгиб ствола с деформацией междоузлия. Изгиб происходит в процессе роста верхушки побега и в пределах его фронтальной плоскости. С приостановкой роста изгиб ствола не связан. Причиной может быть несоответствие скоростей роста двух сторон верхушки. Встречается редко.
1—10(4). Поворот плоскости колонии вокруг собственной оси. Местом поворота является тонкая перетяжка. Поворот может быть на самый различный угол, вплоть до 90°. Возникает после приостановки роста. Часто сопровождается формированием косой перетяжки (рис. 3, 10, к.п) после начала следующего периода роста. В норме косая перетяжка образуется только на зачатке основного побега при его образовании на гидроризе. Встречается периодически.
1—20 (23). Поочередное расположение гидрантов. Морфотип представляет собой смещение гидрантов одного междоузлия по отношению друг к другу по вертикали. Чаще всего первоначальное смещение незначительно, однако. если несколько пар гидрантов подряд отделены друг от друга толстыми перетяжками, то в каждой последовательной паре смещение увеличивается. После образования тонкой перетяжки смещение исчезает. Самое распространенное отклонение от канонического строения Dynamena pumila. Среди всех аномалий на ее долю может приходиться от 1/3 до 1/2 случаев, что составляет 0.6—0.7% от всех нормальных междоузлии.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--