Статья: Незримое одеяние голых тварей

Влияние зимозана (0.2 мг/мл), стимулятора фагоцитоза, на хемилюминесценцию губки Sycon sp.

Изменение хемилюминесценции актинии Aiptasia pulchella после добавления азида натрия (1 мМ) - ингибитора гемсодержащих ферментов.
Полученные результаты позволили нам предположить, что по крайней мере механизмы генерации активных форм кислорода наружной поверхностью губок и белыми кровяными тельцами сходны. Каким образом организовано производство АФК актиний, оставалось до конца не понятным. Чтобы все-таки выяснить это, мы решили идентифицировать ферменты, образующие АФК, непосредственно измеряя их активность. Так как у актиний генерация супероксида наружной поверхностью не была подтверждена методом ЭПР, мы решили, что скорее всего это не NADPH-оксидаза. “Подозрение”, таким образом, пало на миелопероксидазу и двойную оксидазу.
Первый фермент проявляет две активности: пероксидазную (способность окислять разные субстраты с помощью Н2O2) и хлорирующую. В измельченных тканях (гомогенатах) актиний нам удалось обнаружить только первую. Следовательно, мы имели дело с некой пероксидазой, не имеющей хлорирующей активности. Мы склоняемся к тому, что это может быть двойная оксидаза [10].
Поскольку ксантиноксидаза - тоже кальций-зависимый фермент [1] и теоретически могла бы активироваться иономицином, мы решили выяснить ее причастность к генерации АФК у актиний и губок. Ни у тех, ни у других мы не обнаружили ксантиноксидазную активность; по крайней мере, используемый нами метод не выявил каких-либо ее следов [10].
Теперь необходимо было убедиться в том, что генерация АФК морскими беспозвоночными представляет собой активный процесс, а не пассивную утечку вследствие слабой активности каталазы, как происходит у пресноводной рыбы пецилии [9]. Поэтому мы измеряли активности основных антиоксидантных ферментов - супероксиддисмутазы (СОД) и каталазы в гомогенатах актиний и губок. Подобно нейтрофилам и макрофагам, эти животные обладают антиоксидантными ферментами, которые защищают их от повреждающего действия АФК. Однако с генерированием АФК наружными поверхностями актиний и губок такая защита не сбалансирована. У этих беспозвоночных животных активности таких ферментов довольно высоки в сравнении с теми, которые характерны для “профессионально” генерирующих АФК клеток млекопитающего - макрофагов и нейтрофилов морской свинки. Следовательно, можно было исключить пассивную утечку АФК в качестве единственного способа их удаления (таблица) [10].

Итак, механизмы генерации активных форм кислорода наружными поверхностями водных организмов в принципе те же, что слизистыми оболочками, клетками крови, лимфой и спермой высших животных, в том числе человека. Сходство механизмов позволяет предполагать и сходство функций: вероятно, оболочка из кислородных радикалов и Н2O2 защищает “голых” обитателей водоемов от патогенных микроорганизмов. Обладатели же кутикулы, раковины, наружного скелета или ороговевающего эпидермиса не нуждаются в дополнительной защите наружной поверхности посредством АФК. У таких организмов эти формы служат лишь для поддержания постоянного состава внутренней среды (гомеостаза) и для защиты открытых участков тела, непосредственно граничащих с внешней средой (например, жабр у пецилии [9]).
Голые твари, одетые в незримую “одежду” из АФК, могут приносить пользу не только самим себе, но и всему водному биоценозу в целом: генерируемые ими активные формы кислорода освежают воду, препятствуя развитию гнилостной микрофлоры. Для нее, возможно, АФК служат неким “опознавательным знаком”, позволяющим отличать живые организмы от мертвых и не поедать живых - иначе она рисковала бы остаться в конце концов без питания.
Вероятно, такие отношения между животными и бактериями-сапрофитами, которые расплодились в изобиловавшем органическими остатками теплом первичном океане, сложились еще на заре эволюции органического мира, в докембрии. Для большинства животных, по-видимому, этот способ оказался слишком энергоемким (возможно, из-за повышения концентрации растворенного в океанической воде кислорода - продукта оксигенного фотосинтеза древних синезеленых водорослей), и они “прикрыли” свою поверхность осязаемым наружным скелетом из хитина, известняка или целлюлозы. Результатом этого стала так называемая “кембрийская скелетная революция”. До нее же, как известно, все найденные в ископаемом виде животные были голыми. Для тех, кто дерзнул после этой “революции” остаться нагим, вырабатываемые всей поверхностью тела АФК служили и служат невидимым покровом, защищающим от болезнетворных и гнилостных бактерий.
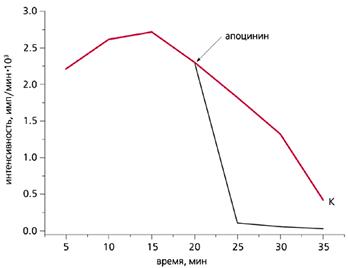
Влияние апоцинина (25 мкМ) на хемилюминесценцию губки Sycon sp.
Этот селективный ингибитор NADPH-оксидазы быстро гасит свечение губки.
В процессе эволюции анатомия живых организмов усложнялась. В слабо упорядоченных сгустках клеток, каковые фактически представляют собой губки, дифференцировались вначале настоящие ткани, затем - органы. Возникали приспособления, позволяющие выходить из водной среды на сушу. Но принципы организации неспецифической защиты организма оставались прежними: участки тела, непосредственно граничащие с внешней средой и не имеющие плотных покровов, оказались защищены невидимым облаком из АФК. У высших позвоночных и человека это желудочно-кишечный тракт, дыхательные пути, гениталии, т.е. органы, которые открыты для внешней среды, но как бы “ввернуты” внутрь тела. Об их защите природа позаботилась, снабдив их способностью генерировать АФК в нужный момент и в необходимом количестве. Интересно, что и нервная ткань, некогда в ходе эволюции граничившая с внешней средой, но у высших организмов “провалившаяся” внутрь тела по ходу индивидуального развития, также сохранила способность к генерации АФК. Так что невидимое одеяние, защищавшее в глубокой древности голых обитателей океана, у одних организмов сохранилось в том же виде и поныне, у других - только в отдельных тканях и органах.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Проект 05-04-49316.
Список литературы
1. Гордеева А.В., Звягильская Р.А., Лабас Ю.А. // Биохимия. 2003. Т.68. №10. С.1318-1322.
2. Geiszt M. // Cardiovasc. Res. 2006. V.71. №2. P.289-299.
3. Arnold J. // Биохимия. 2004. Т.69. №1. С.8-15.
4. Ris-Stalpers C. // Antioxid Redox Signal. 2006. V.8. №9-10. P.1563-1572.
5. Park J. -B. // Exp. Mol. Med. 2003. V.35. №5. P.325-335.
6. Лабас Ю.А., Гордеева А.В. Как возникла биолюминесценция? // Материалы международной конференции “Водные экосистемы и организмы - 4”.Ред. В.Д.Федоров, С.А.Остроумов. М., 2003. С.76-80.
7. Гордеева А.В., Лабас Ю.А. // Цитология. 2003. Т.45. №3. С.284-289.
8. Peskin A.V., Labas Y.A., Tikhonov A.N. // FEBS Lett. 1998. V.434. №1-2. P.201-204.
9. Wilhelm-Filho D., Gonzalez-Flecha B., Boveris A. // Braz. J. Med. Biol. Res. 1994. V.27. №12. P.2879-2882.
10. Гордеева А.В., Наглер Л.Г., Лабас Ю.А. // Журн. эволюционной биохимии и физиологии. 2006. Т.42. №3. С.201-207.