Учебное пособие: Молекулярные механизмы регуляции клеточного цикла
Клеточный цикл – это период жизни клетки от одного деления до другого или от деления до смерти. Клеточный цикл состоит из интерфазы (период вне деления) и самого клеточного деления.
В конце G1 периода принято различать специальный момент, называемый R‑точкой (точка рестрикции, R‑пункт), после которого клетка обязательно в течение нескольких часов (обычно 1–2) вступает в S период. Период времени между R‑точкой и началом S периода можно рассматривать в качестве подготовительного для перехода в S период.
Самый главный процесс, который идет в S периоде, – это удвоение или редупликация ДНК. Все остальные реакции, происходящие в это время в клетке, направлены на обеспечение синтеза ДНК. К таким вспомогательным процессам можно отнести синтез гистоновых белков, синтез ферментов, регулирующих и обеспечивающих синтез нуклеотидов и образование новых нитей ДНК.
Прохождение клетки по всем периодам клеточного цикла строго контролируется. При движении клеток по клеточному циклу в них появляются и исчезают, активируются и ингибируются специальные регуляторные молекулы, которые обеспечивают: 1) прохождение клетки по определенному периоду клеточного цикла и 2 переход из одного периода в другой. Причем прохождение по каждому периоду, а также переход из одного периода в другой контролируется различными веществами. Сейчас мы попробуем выяснить, что же это за вещества и что они делают.
Общая ситуация выгладит так. В клетке постоянно присутствуют специальные белки-ферменты, которые путем фосфорилирования других белков (по остаткам серина, тирозина или треонина в полипептидной цепи), регулируют активность генов, ответственных за прохождение клетки по тому или иному периоду клеточного цикла. Эти белки-ферменты называются циклин-зависимыми протеинкиназами (cdc). Имеется несколько их разновидностей, но они все обладают сходными свойствами. Хотя количество этих циклин-зависимых протеинкиназ может варьировать в различных периодах клеточного цикла, они присутствуют в клетке постоянно, независимо от периода клеточного цикла, то есть они имеются в избытке. Другими словами, их синтез или количество не лимитирует или не регулирует прохождение клеток по клеточному циклу. Однако при патологии, если синтез их нарушен, снижено их количество или имеются мутантные формы с измененными свойствами, то это, конечно же, может повлиять на течение клеточного цикла.
Почему же такие циклин-зависимые протеинкиназы сами не могут регулировать прохождение клеток по периодам клеточного цикла. Оказывается, что они находятся в клетках в неактивном состоянии, а для того чтобы они активировались и начали работать, необходимы специальные активаторы. Ими являются циклины. Их также много разных типов, но они присутствуют в клетках не постоянно: то появляются, то исчезают. В разные фазы клеточного цикла образуются разные циклины, которые связываясь с Cdk образуют различные Cdk‑циклиновые комплексы. Эти комплексы регулируют разные фазы клеточного цикла и поэтому называются G1-, G1/S-, S- и М-Cdk (рис. из моих рис. циклины). Так, например, прохождение клетки по G1 периоду клеточного цикла обеспечивает комплекс циклин-зависимой протеинкиназы‑2 (cdk2) и циклина D1, циклин-зависимой протеинкиназы‑5 (cdk5) и циклина D3. Прохождение через специальную точку рестрикции (R‑пункт) периода G1 контролирует комплекс cdc2 и циклина С. Переход клетки из G1 периода клеточного цикла в S период контролирует комплекс cdk2 и циклина Е. Для перехода клетки из S периода в G2 период необходим комплекс cdk2 и циклин А. Циклин-зависимая протеинкиназа‑2 (cdc2) и циклин В участвуют в переходе клетки из G2 периода в митоз (М период). Циклин H в соединении с cdk7 необходим для фосфорилирования и активации cdc2 в комплексе с циклином В.
| РЕГУЛЯЦИЯ КЛЕТОЧНОГО ЦИКЛА | |
| G1 период | cdk2 + циклин D1 cdk5 и циклин D3 |
| R‑пункт периода G1 | cdc2 + циклин С |
| переход из G1 в S период | cdk2 + цикли Е |
| переход из S в G2 период | cdk2 + циклин А |
| переход из G2 периода в митоз (М период) | cdc2 + циклин В |
| циклин H + cdk7 необходим для фосфорилирования и активациии cdc2 в комплексе с циклином В | |
Циклины – новый класс белков, открытый Тимом Хантом, которые играют ключевую роль в управлении делением клеток. Название «циклины» появилось из-за того, что концентрация белков этого класса изменяется периодически в соответствии со стадиями клеточного цикла (например, падает перед началом деления клетки).
Первый циклин был обнаружен Хантом в начале 1980-х годов, во время опытов с икрой лягушек и морских ежей. Позднее циклины были найдены и в других живых существах.
Оказалось, что эти белки мало изменились в ходе эволюции, как и механизм управления клеточным циклом, который дошел от простых дрожжевых клеток до человека в «законсервированном» виде.
Тимоти Хант (R. Timothy Hunt) вместе с соотечественником-англичанином Полом Нерсом (Paul M. Nurse) и американцем Лиландом Хартуэллом (Leland H. Hartwell) в 2001 году получили нобелевскую премию по физиологии и медицине за открытие генетических и молекулярных механизмов регуляции клеточного цикла – процесса, который имеет важнейшее значение для роста, развития и самого существования живых организмов
Контрольные точки клеточного цикла
1. Точка выхода из G1‑фазы, называемая Старт – у млекопитающих и точкой рестрикции у дрожжей. После перехода через точку рестрикции R в конце G1 наступление S становится необратимым, т.е. запускаются процессы ведущие к следующему делению клетки.
2. Точка S – проверка точности репликации.
3. Точка G2/M‑перехода – проверка завершения репликации.
4. Переход от метафазы к анафазе митоза.
Регуляция репликации
Перед началом репликации Sc ORC‑комплекс (origin recognition complex) садится на ori – точку начала репликации. Cdc6 представлен во всем клеточном цикле, но его концентрация возрастает вначале G1, где он связывается c ОRC комплексом, к которому затем присоединяются Mcm белки с образованием pre-replicative complex (pre-RC). После сборки pre-RC клетка готова к репликации.
Для инициации репликации S-Cdk соединяется с протеинкиназой (?), которая фосфорилирует pre-RC. При этом Cdc6 диссоциирует от ОRC после начала репликации и фосфорилируется, после чего убиквитинируется SCF и деградирует. Изменения в pre-RC препятствуют повторному запуску репликации. S-Cdk так же фосфорилирует некоторые Mcm белковые комплексы, что запускает их экспорт из ядра. Последующая дефосфориляция белков вновь запустит процесс образования pre-RC.
Циклины – активаторы Cdk. Циклины, так же как и Cdk вовлечены в различные, помимо контроля клеточного цикла, процессы. Циклины разделяются на 4 класса в зависимости от времени действия в клеточном цикле: G1/S, S, M и G1 циклины.
G1/S циклины (Cln1 и Cln2 у S. cerevisiae, циклин E у позвоночных) достигает максимальной концентрации в поздней G1‑фазе и падает в S‑фазе.
G1/S cyclin–Cdk комплекс запускает начало репликации ДНК выключая различные системы подавляющие S-phase Cdk в G1‑фазе G1/S циклины также инициируют дупликацию центросом у позвоночных, образование веретенного тела у дрожжей. Падение уровня G1/S сопровождается увеличением концентрации S циклинов (Clb5, Clb6 у Sc и циклин A у позвоночных), который образует S циклин-Cdk комплекс который напрямую стимулирует ДНК репликацию. Уровень S циклина остается высоким в течении всей S, G2‑фаз и начала митоза, где помогает началу митозу в некоторых клетках.
М-циклины (Clb1,2,3 и 4 у Sc, циклин B у позвоночных) появляется последним. Его концентрация увеличивается, когда клетка переходит к митозу и достигает максимума в метафазе. М-циклин-Cdk‑комплекс включает сборку веретена деления и выравнивание сестринских хроматид. Его разрушение в анафазе приводит к выходу из митоза и цитокиезу. G1 циклины (Cln3 у Sc и циклин D у позвоночных) помогает координировать клеточный рост с входом в новый клеточный цикл. Они необычны, так как их концентрация не меняется от фазы клеточного цикла, а меняется в ответ на внешние регуляторные сигналы роста.
Программируемая клеточная гибель
В 1972 г. Керр с соавт. опубликовали статью, в которой авторы представили морфологические доказательства существования отличающегося от некроза особого вида гибели клеток, которую они назвали «апоптоз». Авторы сообщили, что структурные изменения клеток при апоптозе проходят две стадии:
1-я – образование апоптозных тел,
2-я – их фагоцитоз и разрушение другими клетками.
Причины гибели, процессы морфологического и биохимического характера развития клеточной смерти могут быть различными. Но все же их можно четко разделить на две категории:
1. Некроз (от греч. пеkrosis– омертвление) и
2. Апоптоз (от греч. корней, означающих «отпадение» или «распадение»), который часто называют программируемой клеточной смертью (ПКС) или даже клеточным самоубийством (рис. 354).
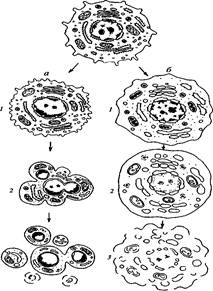
Два пути клеточной гибели
а – апоптоз (профаммированная клеточная смерть): / – специфическое сжатие клетки и конденсация хроматина, 2 – фрагментация ядра, 3 – фрагментация тела клетки на ряд апоптических телец; б – некроз: / – набухание клетки, вакуолярных компонентов, конденсация хроматина (кариорексис), 2 – дальнейшее набухание мембранных органоидов, лизис хроматина ядра (кариолизис), 3 – разрыв мембранных компонентов клетки – лизис клетки
НЕКРОЗ
Н. является наиболее частой неспецефической формой гибели клеток. Он может быть вызван тяжелыми повреждениями клетки в результате прямой травмы, радиации, влияния токсических агентов, вследствие гипоксии, лизиса клетки, опосредованного комплементом и т.д.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--