Дипломная работа: Нарушение экспрессии D-глюкуронил С5-эпимеразы как возможная причина изменения структуры протеогликанов в опухоли молочной железы человека
К тому же цепи ГАГ значительно превосходят по длине углеводные цепи гликопротеинов. Например, цепь ГАГ с молекулярным весом 20 кДа содержит около 50 остатков сахаров, в то время как типичный N-гликан содержит 10-12 остатков.
Протеогликаны могут нести на своем коровом белке помимо ГАГ-цепей также короткие нерегулярные олигосахариды. При этом свойства сложной молекулы определяются в основном типом ГАГ-цепи, хотя олигосахариды могут также влиять на биологическую активность.
Если к гликопротеину присоединена хотя бы одна цепь ГАГ, он является протеогликаном.
1.3 Классификация протеогликанов
Классифицируют протеогликаны по типу доминантных цепей ГАГ, присоедненных к коровому белку.
1.4 Строение и классификация гликозаминогликанов
Гликозаминогликаны представляют собой линейные гетерополисахариды, составленные из повторяющихся дисахаридных единиц, каждая из которых состоит из гексозамина ( D-глюкозамина - GlcN или D-галактозамина - GalN), соединенного с гексуроновой кислотой (D-глюкуроновой - GlcUA, или L-идуроновой - IdоUA), присутствующей во всех ГАГ, за исключением кератансульфата, где она замещена остатками галактозы (Gal).
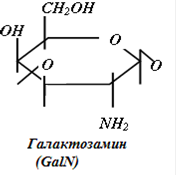
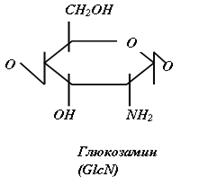
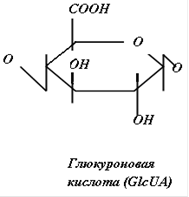
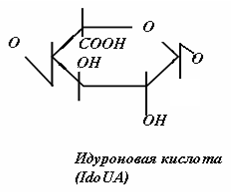
Дисахаридные единицы могут быть N - и O-сульфатированы и/или ацетилированы. Степень сульфатирования ГАГ намного выше, чем других макромолекул и достигает 3-4 сульфатных групп на дисахарид.
Благодаря сульфатным и карбоксильным группам ГАГ имеют очень высокую плотность отрицательного заряда, что во многом определяет их биологические свойства и регулирует их взаимодействие с другими молекулами.
В зависимости от структуры дисахарида, а также степени сульфатирования, все ГАГ разделяют на несколько классов: гиалуроновая кислота (ГК), хондроитинсульфаты (ХС), дерматансульфаты (ДС), гепарансульфаты (ГС), гепарин (Ге), кератансульфаты (КС) (таблица 1 и рис.2). Зачастую на одном белковом коре находятся разные типы ГАГ-цепей.
Таблица 1. Классы гликозаминогликанов (ГАГ)
|
Класс ГАГ |
Мол. Вес, кДа |
Повторяющаяся дисахаридная единица |
Сульфогруппа (O-, N-связанная) |
Количество сульфо-групп на 1 дисахарид | Другие сахара, встречаю-щиеся в связующем участке |
| Гиалуроновая кислота, ГК | 1-8000 |
D-GlcUA, D-GlcN | - | - | - |
|
Хондроитин- сульфат, ХС | 10-50 |
D-GlcUA, D-GalN | O- | 0,1-1,3 | D-Gal, D-Xyl |
| Дерматан-сульфат, ДС | 10-40 |
D-GlcUA/ L-IdоUA, D-GalN | O- | 1-2,5 | D-Gal, D-Xyl |
| Гепаран-сульфат, ГС | 10-40 | D-GlcUA/L-IdоUA, D-GlcN | O-, N- | 0,4-2,0 | D-Gal, D-Xyl |
| Гепарин, Ге | 5-25 | D-GlcUA/L-IdоUA, D-GlcN | O-, N- | 1,5-3,0 | D-Gal, D-Xyl |
| Кератан - сульфат I, KCI | 5-25 | D-Gal, D-GlcN | O- |
0,9-1,8 | D-Man, L-Fuc, SA |
| Кератан - сульфат II, KCII | 5-15 | D-Gal, D-GlcN | N- | 0,9-1,8 | D-GalN, SA |
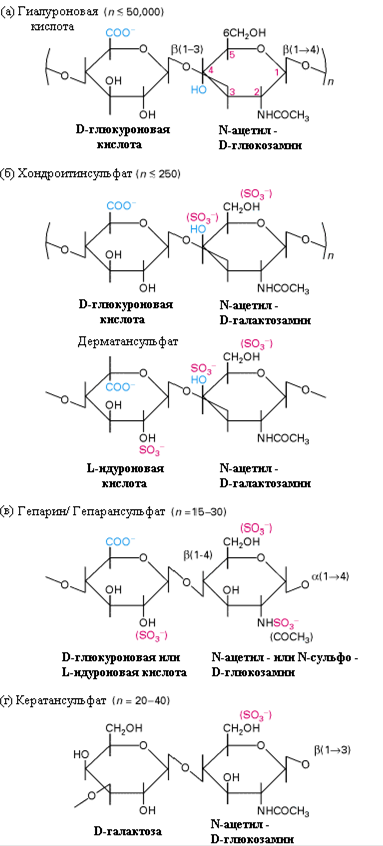
Рис.2. Структура дисахаридов, образующих цепи ГАГ различных классов
1.5 Присоединение цепей ГАГ к коровому белку
Цепи большинства ГАГ присоединяются к остатку серина корового белка через тетрасахаридный линкер: - Xyl-Gal-Gal-GlcUA-, далее следуют соответствующие чередующиеся дисахаридные единицы (рис.3). ГАГ-цепи присоединяются к остаткам серина определенных ГАГ-акцепторных сайтов на коровом белке: "-A-Ser-Gly-X-Gly-", где А - кислая аминокислота, а Х - точно не определена [1]. Дипептид Ser-Gly в последовательности акцепторного сайта белка является основным требованием для узнавания ферментом ксилозилтрансферазой, переносящим остаток ксилозы на серин [2].