Курсовая работа: Гинецей растений
нуцеллус, ядро семязачатка;
фуникулюс, семяножка, с помощью которой семязачаток крепится к плаценте;
интегументы, покровы семязачатка, образующие на верхушке нуцеллуса канал;
микропиле, пыльцевход;
халаза, базальная часть семязачатка, где сливаются нуцеллус и интегументы;
рубчик - место присоединения семязачатка к семяножке.
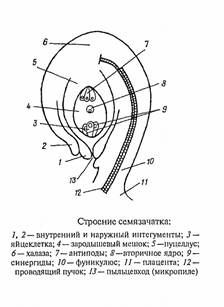
Рис.4 Строение семязачатка
Типы семязачатка:
ортотропный - прямой, фуникулюс и микропиле расположены на противоположных концах оси семязачатка (гречишные, ореховые);
анатропный (обратный) - нуцеллус повернут по отношению к прямой оси семязачатка на 180, вследствие чего микропиле и фуникулюс расположены рядом (Покрытосеменные)
гемитропный (полуповернутый) - семязачаток повернут на 90, вследствие чего микропиле и нуцеллус расположены по отошению к фуникулюсу под углом 90 (первоцветные, норичные).
кампилотропный (односторонне изогнутый) - нуцеллус изогнут односторонне микропилярным концом, соответсвенно микропиле и фуникулюс расположены рядом (бобовые, мальвовые)
амфитропный (двусторонне изогнутый) - нуцеллус изогнут двусторонне в виде подковы, при этом микропиле и фуникулюс располагаются рядом (тутовые, ладанниковые).
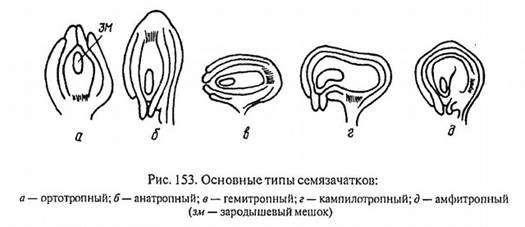
Рис.5 Основные типы семязачатков
6. Развитие семязачатка
Мегаспорогенез протекает в женской репродуктивной сфере - в гинецее. Морфологически гинецей представлен пестиком (или пестиками). В состав пестика входят: рыльце, столбик и завязь. Внутри завязи содержатся семязачатки (один или несколько). Внутреннее содержимое семязачатка представляет собой нуцеллус. Покровы семязачатка образованы двойным или одиночным интегументом. В нуцеллусе семязачатка имеется одна археспориальная клетка (2n), способная делиться путем мейоза (у ив и некоторых других растений археспорий многоклеточный). В результате мейоза из археспориальной клетки (материнской клетки мегаспор) образуется четыре гаплоидные мегаспоры (n). Вскоре три из них отмирают, а одна увеличивается в размерах и трижды делится путем митозов. В результате образуется восьмиядерный зародышевый мешок (женский гаметофит). Три ядра вместе с прилегающей цитоплазмой образуют клетки-антиподы, два ядра - одно центральное диплоидное ядро; два ядра - две клетки-синергиды; одно ядро становится ядром яйцеклетки.
Когда к семязачатку приближается пыльцевая трубка, он это заранее "чувствует" и готовится ее встретить. Клетки-спутницы начинают выделять слизистые вещества. Тем временем пыльцевая трубка растет, преодолевая сопротивление клеток стенок завязи. Наконец, она достигает микропиле. Происходит "драматический" процесс: пыльцевая трубка протыкает (и при этом убивает) одну из клеток-спутниц. Оба спермия покидают пыльцевую трубку. Участь вегетативной клетки пыльцевого зерна незавидна, она вскоре погибнет. Наблюдать этот процесс очень трудно, но еще труднее понять, что происходит при оплодотворении.
В августе 1898 года, когда еще не было известно о двойном оплодотворении, в Киеве состоялся десятый Съезд русских естествоиспытателей и врачей. Профессор Сергей Гаврилович Навашин сделал на нем важное сообщение: оба спермия, содержащиеся в пыльцевом зерне, необходимы для нормального развития семян двух видов из семейства Лилейные (Liliaceae): лилии (Lilium martagon) и рябчика (Fritillaria tenella). Почему Навашин выбрал именно эти растения? Наверное, потому, что спермии и зародышевый мешок у них крупные, их легко рассматривать в микроскоп. Навашев впервые заявил, что в процессе оплодотворения участвуют два спермия, сливающиеся с двумя (!) клетками зародышевого мешка. Но предоставим слово самому автору открытия.
Каждый раз, когда пыльцевая трубка наблюдалась в соприкосновении с зародышевым мешком, оба мужские половые ядра также наблюдались в содержимом зародышевого мешка. Мужские ядра лежат сначала близко друг от друга...
Мужские ядра затем отделяются друг от друга, причем одно проникает к яйцеклетке, а другое тесно прикладывается к одному из еще не слившихся в это время полярных ядер, именно к сестринскому ядру яйцеклетки...
В то время, как мужское ядро все более и более тесно прилегает к ядру яйца, полярное ядро, копулирующее с другим мужским ядром, направляется навстречу к другому полярному ядру, с которым встречается в середине зародышевого мешка...
Только после прохождения профазы деления ядра сливаются..."
Чтобы объяснение было понятнее, Сергей Гаврилович сделал к рисунки, очень понравившиеся столичным ботаникам. Они отвезли их, чтобы показать на заседании Петербургской Академии.
Почему же этому выступлению ученые придали такое большое значение? Считалось, что оплодотворение у растений происходит точно так же как у животных. Один спермий и одна яйцеклетка должны давать зиготу, из которой развивается новое растение. Не то чтобы ботаники не видели двух спермиев, движущихся в пыльцевой трубке к зародышевому мешку. Считалось, что это - какая-то ненормальность, "типичное" растение должно иметь не два, а один спермий, "типичное" оплодотворение должно быть одинаковым и у растении и у животных. Оказалось, что эти представления были неправильными. Оплодотворение цветковых растений совершенно не похоже на оплодотворение не только животных, но и других растений. Процесс оплодотворения с помощью двух спермиев получил название двойного оплодотворения. Сразу же после открытия Навашина ученые бросились к своим препаратам. Оказалось, что многие уже видели двойное оплодотворение, но считали его уродливым процессом, якобы не дающим нормальных семян. Ученые стали присылать Сергею Гавриловичу поздравления. А один ученый даже подарил свои старые препараты, на которых было хорошо видно двойное оплодотворение.
Итак, С.Г. Навашин открыл удивительное явление. Зачем происходит оплодотворение яйцеклетки было понятно: чтобы получилась зигота, а из нее - новое растение. Но зачем же оплодотворять центральную клетку зародышевого мешка? Оказалось что именно из этой клетки развивается питательная ткань семени - эндосперм. Навашин предположил, что без мужского ядра эндосперм образоваться не может. Как же проверить это предположение? Ведь из пыльцевой трудки невозможно вытащить один из спермиев, не нарушая процесс оплодотворения.
Сергей Гаврилович решил исследовать растения, у которых нет эндосперма. Вдруг у них нарушено оплодотворение центральной клетки? Такие растения отыскались в семействе Орхидных (Orchidaceae). Семена орхидных очень мелкие, и даже прорости самостоятельно не могут (а прорастают только при помощи грибов базидиомицетов, образуя микоризу). В пыльцевой трубке орхидных Навашин увидел два спермия. Один из них оплодотворял яйцеклетку, а второй "пробовал" оплодотворить центральную клетку с двумя полярными ядрами. Но в центральной клетке ядра не сливались друг с другом! Процесс оплодотворения был нарушен и, естественно, эндосперм не мог образоваться. В дальнейших работах ученому удалось показать, что у подсолнечника (Helianthus annuus) и некоторых других растений оплодотворение - двойное. После того, как Навашин нашел двойное оплодотворение у самых разных растений, он сделал вывод, что двойное оплодотворение свойственно всем цветковым растениям.
Взглянем на строение семязачатка после двойного оплодотворения. Клетки обоих его покровов как имели два набора хромосом, так их и сохранили. Оба набора в этих клетках принадлежат материнскому растению. Нуцеллус также несет два материнских наборов хромосом. В зародышевом мешке синергиды погибли, а у антиподов по одному набору хромосом. Эти наборы - тоже материнские. Зигота, образовавшаяся слиянием яйцеклетки и спермия, несет два набора хромосом: один от отцовского, а другой от материнского растения. Самое интересное, что центральная клетка имеет три набора хромосом: один от отцовского растения, и два - от материнского.
После двойного оплодотворения начинается несколько процессов: первичное ядро эндосперма делится, образуя эндосперм, зигота развивается в зародыш, интегументы превращаются в семенную кожуру, а стенка завязи и связанные с ней структуры формируют плод. На ранних стадиях развития последовательность клеточных делений у зародышей двудольных и однодольных сходна; в обоих случаях формируются сферические тела. Позже появляется различие: зародыш двудольных имеет две семядоли, а однодольных - только одну. В семенах некоторых групп покрытосеменных нуцеллус развивается в запасную ткань, называемую перистермом. Некоторые семена содержат и эндосперм и перистерм, так у свеклы (Belta). Однако у многих двудольных и некоторых однодольных вся запасная ткань или большая ее часть поглощается развивающимся зародышем еще до того, как семя впадет в состояние покоя (у гороха, у бобов и др.). Зародыши в таких семенах обычно развивают заполненные питательными веществами мясистые семядоли. Основные питательные вещества, запасаемые в семенах, - углеводы, белки, липиды. Семена голосеменных и покрытосеменных различаются происхождением этих веществ. У первых их дает женский гаметофит, у вторых - эндосперм, не являющийся ни гаметофитной, ни спорофитной тканью.