Курсовая работа: Гинецей растений
Так происходит развитие гаметофита, оплодотворение и формирование семени у "типичного" цветкового растения, однако мир растений очень разнообразен и у множества покрытосеменных развитие проходит иначе, однако не менее интересно и удивительно. Все эти способы развития помогают каждому растению поддерживать свою стратегию в борьбе за существование; преимущество двойного оплодотворения не ограничивается тем, что растения формируют питательные запасы в семязачатке только при условии оплодотворения (не "растрачивая" силы на формирование питательных запасов в семязачатке, который никогда не будет оплодотворен, как это часто происходит у голосеменных), эти различия в развитии, являющиеся следствием приспособленности растений к окружающим их условиям, и создают то удивительное многообразие форм морфологического и физиологического строения, которое мы и наблюдаем у цветковых.
7. Зародышевый мешок и его развитие
Женский гаметофит цветковых растений обычно называется зародышевым мешком. Возникновение этого названия восходит к тем временам, когда еще не была ясна морфологическая природа чрезвычайно своеобразного женского гаметофита цветковых растений. В настоящее время оба термина употребляют как синонимы.
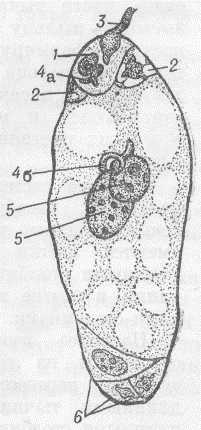
Рис.6. Зародышевый мешок (женский заросток) покрытосемянного (цветкового) растения и двойное оплодотворение: 1 - яйцеклетка; 2 - клетки-помощницы; 3 - вскрывшаяся пыльцевая трубка. Один из спермиев (4а) сливается с яйцеклеткой; 5 - полярные ядра, сливающиеся со вторым спермием (46); 6 - три клетки в нижней части зародышевого мешка (антиподы).
У большинства цветковых растений (вероятно, не менее чем у 80%) начало женскому гаметофиту дает одна из четырех потенциальных мегаспор тетрады путем трех последовательных митотических делений. Этот тип образования женского гаметофита называется моноспорическим. Его развитие происходит следующим образом. В результате первого деления ядра функционирующей мегаспоры образуются два ядра, которые расходятся к полюсам (микропилярному и халазальному) сильно удлиняющейся мегаспоры, а между ними образуется крупная вакуоль. Далее каждое из этих двух ядер делится синхронно еще два раза и в результате у каждого полюса образуются по четыре ядра. Это восьмиядерная стадия развития женского гаметофита. От каждой из двух полярных четверок входит по одному ядру в центральную часть зародышевого мешка. Эти так называемые полярные ядра сближаются в центральной части и сливаются или сразу же, или только перед оплодотворением, а иногда и вовсе не сливаются. В результате слияния полярных ядер образуется ядро центральной клетки женского гаметофита, называемое часто вторичным ядром зародышевого мешка.
В дальнейшем (в процессе последнего митоза) возникают клеточные перегородки и свободноядерная стадия развития женского гаметофита переходит в клеточную стадию. У микропилярного полюса образуются три клетки яйцевого аппарата, у противоположного (халазального) - три клетки, называемые антиподами, а между этими двумя группами клеток - центральная клетка (содержащая два свободных полярных ядра или одно вторичное ядро). Три клетки яйцевого аппарата дифференцируются на одну яйцеклетку и две клетки, называемые синергидами (от греч. synergos - вместе действующие) или вспомогательными клетками. Таково строение так называемого нормального моноспорического женского гаметофита.
Нормальный моноспорический женский гаметофит не производит впечатления окончательно отработанной модели, в которой экономичность достигла такого же предела, как в случае мужского гаметофита. То, что предел здесь не достигнут, доказывается следующим. В процессе эволюции этого основного для цветковых растений типа женского гаметофита возникли зародышевые мешки других типов, образуемые двумя или даже четырьмя мегаспорами. Произошло сокращение митотических делений до двух или одного, а также изменилось распределение ядер. Разные сочетания этих изменений обусловили возникновение нескольких типов женского гаметофита, которые отличаются как числом ядер (4, 8, 16), числом клеточных групп и полярных ядер (1, 2, 4, 7-14), так и числом клеток в группах (например, яйцевой аппарат может состоять из 1, 2, 3, 5 и 7 клеток), а также другими признаками.
Основной особенностью моноспорического типа образования женского гаметофита является последовательное заложение клеточных перегородок при мейотических делениях и образование обособленных мегаспор. Совершенно иной характер имеет мегаспорогенез, предшествующий образованию биспорического (двухспорового) и тетраспорического (четырехспорового) гаметофитов. Изменения в мегаспорогенезе заключаются в подавлении заложения клеточной перегородки при мейотическом делении, вследствие чего вместо тетрад мегаспор возникают неклеточные образования, называемые ценоцитами (от греч. koinos - общий и kytos - сосуд, клетка), которые и развиваются далее в женские гаметофиты. Здесь возможны два варианта.
В первом случае образование клеточных перегородок подавляется только при втором делении мейоза. Поэтому оба ядра, вошедшие в состав жизнеспособной клетки диады, принимают участие в образовании женского гаметофита. Двухъядерная диада - это ценоцит, продукт слияния (вернее, неразделения) двух потенциальных мегаспор. Поскольку каждый такой ценоцит соответствует по происхождению паре необособившихся мегаспор, его рассматривают как двухспоровый ценоцит, а развивающийся из него женский гаметофит - как биспорический (двуспоровый). Биспорический женский гаметофит наблюдается у лука, пролески (Scilla), ландыша, амариллисовых, частуховых и некоторых других растений. Развитие такого женского гаметофита происходит более сокращенным путем и для образования восьмиядерной стадии требуется всего лишь два митотических деления.
Наконец, у некоторых растений, таких, как, пеперомия, ильм, майник, рябчик, тюльпан, лилия, гусиный лук, плюмбаго, адокса и другие, образование клеточных перегородок подавляется при обоих мейотических делениях. Вся материнская клетка мегаспор превращается здесь в один четырехспоровый ценоцит, или тетраценоспору. Четырехспоровый ценоцит соответствует четырем неразделившимся мегаспорам, почему женские гаметофиты этого типа называются тетраспорическими (четырехспоровыми). Для образования восьмиядерной стадии женского гаметофита требуется лишь одно митотическое деление ядер четырехспорового ценоцита.
Согласно И.Д. Романову, в отличие от моноспорического женского гаметофита, би - и тетраспорические называются ценоспорическими.
Женский гаметофит цветковых растений произошел из более примитивного архегониального женского гаметофита далеких голосеменных предков путем выпадения последних стадий развития, ведущих к формированию архегониев. А у ценоспорических типов происходит, как мы видели, выпадение также начальных стадий. Но в то время как эволюция мужского гаметофита достигла кульминационной стадии и в его развитии полностью выпали все возможные начальные и конечные стадии, с женским гаметофитом эволюционный процесс как бы продолжает экспериментировать. Отсюда значительное число моделей женского гаметофита и путей его формирования, на чем здесь нет, однако, возможности останавливаться.
Литература
1. http://ru. wikipedia.org/wiki
2. И.И. Андреева, Л.С. Родман "Ботаника"/ Москва "КолосС" 2002
3. П. Рэйвн, Р. Эверт, С. Айкхорн "Современная ботаника"/ Москва "Мир" 1990
4. Магешвари П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Тахтаджян А.Л., Высшие ра