Курсовая работа: Особенности биологии и экологии Аphroditiformia Баренцева моря
Сем. Aphroditidae (7, 63)
Сем. Polynoidae (93, 539)
Сем. Sigalionidae (17, 156)
Сем. Pholoididae (1, 4)
Сем. Eulepethidae (4, 12)
Сем. Polyodontidae (6, 39)
Надсем. Chrysopetalacea
Сем. Chrysopetalidae (4, 24)
Сем. Palmyridae (1,2)
Надсем. Pisionacea Сем. Pisionidae (4, 14)
Надсемейство Aphroditacea является центральной группой в филогенетическом отношении, обладает большим количеством родов и видов, отличающихся морфологическим разнообразием. Наибольшее в этом надсемействе значение имеет семейство Polynoidae. Надсемейство Chrysopetalacea представляет собой как бы связующее звено с высокоспециализированным семейством Amphinomida . Надсемейство Pisionacea занимает промежуточное положение между Aphroditiformia и Glyceriformia ,причем по личиночному развитию пизиониды ближе всего к чешуйчатым червям.
2. КРАТКИЙ МОРФО-ФУНКЦИОНАЛЬНЫЙ ОЧЕРК APHRODITIFORMIA
2.1 ФОРМА ТЕЛА И РАЗМЕРЫ
Тело чешуйчатых червей, как правило, заметно сплющено дорсовентрально. Спинная сторона, прикрытая спинными чешуйками, или элитрами, несколько выпуклая (сводчатая), а брюшная — более плоская, с медиальным продольным желобком. К переднему и особенно к заднему концам тело немного суживается; на заднем конце тела все сегменты становятся более короткими и узкими. Многие чешуйчатые черви обладают относительно коротким телом, имеют овальную форму; у других же, наоборот, оно сильно вытянутое («червеобразное»). Форма тела связана с образом жизни. Удлиненное тело характерно для червей, глубоко зарывающихся в грунт (многие Sigalionidae), строящих собственные длинные трубки (Polyodontidae), а также для червей, ведущих комменсальный образ жизни (некоторые Polynoidae). У чешуйчатых червей наблюдается вполне определенная тенденция к стабилизации количества сегментов и к укорочению тела, что для них, бесспорно, является эволюционно прогрессивным моментом.
Форма тела чешуйчатых червей, в противоположность многим другим поли-хетам, весьма постоянная, не подверженная сколько-нибудь заметному удлинению и сжиманию при передвижениях, причем она мало изменяется и при фиксации. Некоторые полиноиды с укороченным телом (в основном Lepidonotus) при фиксации сворачиваются в колечко (уже по этой особенности они хорошо распознаются среди прочих полиноид).
Размеры чешуйчатых червей варьируют. Представители семейства Aphroditidae s. str. характеризуются довольно крупными размерами. Так, Aphrodita aculeata, весьма обычная у берегов Европы, нередко имеет в длину около 100 мм, при ширине 40—50 мм (отмечены экземпляры до 180 мм длины), a Laetmonice producta из южного полушария достигает длины 135 мм и ширины до 40— 65 мм, включая параподии со щетинками. В семействе Polynoidae также наблюдаются гиганты. В частности, к ним относится калифорнийская полиноида Hololepida magna длиной 250 мм. Из антарктических районов в коллекциях ЗИН АН СССР имеются экземпляры Eulagisca gigantea, достигающие в длину 210 мм при ширине 60 мм без щетинок и 100 мм со щетинками. В арктических морях одна из наиболее крупных полиноид — Eunoe nodosa — до 100 мм длины и 35 мм ширины. Средняя же длина большинства видов полиноид около 30—40 мм. Наиболее тонкие и длинные полиноиды — Lepidasthenia (до 200 мм длины), характеризующиеся также наибольшим количеством сегментов.
Великанами среди полихет справедливо считаются некоторые Polyodontidae, живущие в очень крупных длинных илистых трубках, в связи, с чем их иногда называют «илистыми сосисками»; так, имеются указания, что они достигают в длину 2 метров при ширине 2 см. Среди сигалионид немало видов до 300 мм длины при ширине 8—9 мм (Sigalion squamatum и др.). Но наряду с такими крупными формами среди чешуйчатых червей есть виды, которые характеризуются крайне небольшими размерами., не превышающими 20—25 мм. При этом длина тела не коррелирует с количеством сегментов. Например, Pholoe minuta с 70 сегментами имеет в длину максимум 20 мм, а у упомянутого выше Laetmonice producta, достигающего 135 мм, количество сегментов не свыше 50; у Eulagisca gigantea и Harmothoe Imbricata количество сегментов не превышает 37—38, в то же время длина особей первого вида иногда превосходит 200 мм, а второго — достигает только 50 мм, обычно около 30 мм.
Наиболее разнообразна окраска элитр. Последние могут иметь пестрый или узорчатый рисунок, непосредственно связанный со скульптурой самих элитр. Исключительно красив бросающийся в глаза рисунок в виде продольных полос на элитрах Hyperhalosydna striata. Уже по этому рисунку удается безошибочно определить вид. Иногда элитры бесцветные и полупрозрачные, — например, у видов Aphrodita, элитры которых прикрыты наружным войлоком из волосовидных щетинок, или у Bylgides sarsi, у которого наличие тонких и нежных элитр связано с его факультативным планктонным образом жизни. У Psammolyce, зарывающегося в песок, элитры, как правило, прочно обклеены мелкими песчинками. Это, по всей видимости, своего рода маскировка, (предохраняющая червей от врагов). Благодаря относительно темной окраске тела, в особенности элитр, соответствующей общему фону субстрата, на котором поселяются полиноиды, последних не всегда удается быстро обнаружить. К этому следует добавить, что элитры крупных ползающих по поверхности грунта полиноид часто покрыты разнообразными эпифитами и различными комменсалами, что еще больше скрывает их на морском дне.
Щетинки большей частью золотистого или светло-янтарного оттенка, могут быть и совершенно прозрачными. Наряду с этим крупные спинные игловидные щетинки Aphrodita почти черного цвета.
2.2 СЕГМЕНТАЦИЯ И ДЕЛЕНИЕ ТЕЛА НА ОТДЕЛЫ
Тело многощетинковых червей поделено на сегменты (членики), которые внешне хорошо распознаются по наличию на каждом из них по одной паре параподий, снабженных обычно различными придатками. Наружная кольчатость тела тесно связана с внутренними септами (диссепиментами),: обособляющими друг от друга отдельные сегменты (метамеры, или сомиты) (рис. 1). В каждом сегменте, как правило, закономерно повторяются наборы внутренних органов; комплекс нервных образований одного сегмента обозначается как невросомит, сплетение кровеносных сосудов — ангиосомит, система мышечных волокон — миосомит и т. д.; большей частью метамерно располагаются также выделительные органы и гонады. У чешуйчатых червей кишечник в каждом сегменте имеет боковые слепые выросты (карманы, или дивертикулы), так что и он у них обладает вполне определенной метамерией (рис. 2).
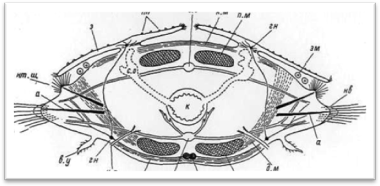
Рис. 1. Поперечный разрез через туловищный сомит (чешуйчатый сегмент).
Таким образом, наружная кольчатость, или наружная метамерия, червей обычно соответствует внутренней метамерии.1
Полная гомономность (однородность) всех туловищных сегментов у многощетинковых червей никогда не наблюдается. Наибольшая гомономность сегментов характерна для бродячих (эррантных) полихет, но и у них она в той или другой степени нарушена. Следует различать гетерономность первичную и вторичную. Первичная гетерономность возникает еще в процессе метаморфоза личинки и связана с образованием ларвальных и постларвальных сегментов, имеющих разное происхождение. Ларвальные сегменты закладываются обычно одновременно на начальной стадии метаморфоза личинок, а лостларвальные сегменты последовательно и постепенно возникают из предпигидиальной эктодермальной зоны роста, причем целом в ларвальных сегментах всегда формируется из мезодермальных полосок путем их разделения на мезодермальные блоки, которые затем становятся полыми.
1 При описании и на рисунках червей сегменты обозначаются римскими цифрами
Число ларвальных сегментов варьирует: у Polynoidae их 8—10, а у Siga-lionidae — 4—6. Большое и изменчивое количество ларвальных сегментов указывает на примитивный тип метаморфоза. Ларвальные сегменты в отличие от постларвальных не образуют половых желез, не имеют целомодуктов и хлора-гогенных клеток. Иногда ларвальные сегменты подвергаются слиянию, образуя перистомиум: например, крупный перистомиум нереид представляет собой продукт слияния трех передних ларвальных сегментов. Вторичная гетерономность относится к постларвальным сегментам, она целиком обусловлена особенностями обитания взрослых червей (адаптивные приспособления).
Количество сегментов у чешуйчатых червей, как и у большинства других полихет, сильно варьирует. Существуют виды, у которых сегментов относительно немного и их число строго постоянное (например, Lepidonotus — всего 26 сегментов), но некоторые виды (многие Lepidasthenia, Arctonoe и др.) состоят из 100 и более сегментов, причем количество последних увеличивается с возрастом и возникновение новых сегментов на заднем конце тела (в предпиги-диальной зоне роста) не прекращается с наступлением половой зрелости. Самые задние сегменты обычно меньше (тоньше и короче) остальных и имеют недоразвитые параподий. Вполне естественно, что черви с большим количеством сегментов одновременно и наиболее длинные. Однако, как уже отмечалось ранее, количество сегментов не всегда строго коррелирует с размерами самих червей. Неопределенно большое количество сегментов у полиноидных червей мы рассматриваем как примитивное состояние, а стабилизацию количества сегментов — как прогрессивный момент. Подсчет сегментов легче всего производить с брюшной стороны, так как спинная сторона червей нередко замаскирована различными придатками, в частности у чешуйчатых червей элитрами. Количество сегментов у представителей афродитид (Aphro- f dita, Laetmonice, Hermonia, Pontogenia) не превышает 50 (колеблется в пределах 32—47). Среди полипоид по числу сегментов выделяется три группы родов: 1) сегментов 25— 30 — Lepidonotus, Hermenia, Euphione, Iphione; 2) сегментов 30—50 — Harmothoe, Lagisca, Eunoe, Antinoe, Gattyana, Bylgides, Eucranta, Malm-grenia, Alentia и др. и 3) сегментов свыше 50 (до 100 и более) — Perolepis, Arctonoe, Lepidasthenia и др. Для большинства сигалионид характерно более 50 сегментов.