Курсовая работа: Особенности биологии и экологии Аphroditiformia Баренцева моря
Помимо основных наружных перетяжек, отделяющих соседние сегменты друг от друга, в пределах одного сегмента преимущественно на его спинной стороне нередко наблюдается добавочная вторичная, или так называемая ложная кольчатость — псевдосегментация (annuli). У рассматриваемых нами чешуйчатых червей она имеет характер небольших метамерно повторяющихся бороздок или складок, к которым приурочены ресничные пояски и поперечные пигментные полосы, придающие некоторым полиноидам весьма пеструю окраску.
При беглом взгляде на чешуйчатых червей нередко создается впечатление, что они обладают гомономной сегментацией, т. е. на всем протяжении тела у них все сегменты однородны. Такое впечатление объясняется тем, что у многих полиноид вся их спинная сторона как бы равномерно прикрыта спинными чешуйками (элитрами). Однако это ложное представление, так как элитры имеются далеко не на всех сегментах. У чешуйчатых червей наблюдается вполне определенное чередование усиковых сегментов, лишенных элитр, и чешуйчатых сегментов, снабженных элитрами. Таким образом, говорить о полной гомономности сегментов у них не приходится.
Многократное повторение двух или трех смежных сегментов, имеющих разное морфологическое строение, называется метамерией 2-го порядка.
Метамерия 2-го порядка, создающая видимость гомономности, имеет место не только у чешуйчатых червей. Например, у некоторых силлид более длинные спинные усики нередко чередуются через сегмент с более короткими, а иногда (Parasphaerosyllis) изменяется поочередно и форма самих усиков.
Метамерия 2-го порядка — это лишь один из примеров наиболее простой дифференциации сегментов. Она представляет собой начальную стадию образования гетерономности. Наиболее гомономными червями являются филло-доциды, отчасти нереиды. У большинства полихет, в особенности у седентарных, ведущих тубикольный образ жизни, процесс возникновения гетерономности идет значительно дальше и видоизмененные сегменты не чередуются между собой, а приурочены к различным участкам тела, выполняющим разные функции. В связи с этим у некоторых полихет резко обособляются передний участок тела — торакс (так называемые грудные сегменты) и задний участок — абдомен (брюшные сегменты), а иногда и дополнительный хвостовой участок. Наиболее сложная гетерономия наблюдается у ряда седентарных полихет, в частности у Chaetopteridae и Sabellariidae, что обусловлено тубикольным образом жизни. У чешуйчатых червей разделения тела на торакс и абдомен нет, однако XXIII сегмент у них все же играет какую-то роль, отделяя переднюю часть тела со строго закономерным расположением элитр через один сегмент от задней, где расположение элитр еще не стабилизировалось.
У всех чешуйчатых червей, как и у остальных полихет, различают три основных отдела тела. Во-первых, передний, или так называемый головной, отдел, слагающийся из простомиума, или головной лопасти, и двух передних сегментов — перистомиального и буккального. Во-вторых, туловищный отдел (метастомиум), состоящий из большого количества повторяющихся сегментов с развитыми параподиями; этот отдел является наиболее крупной частью червя. В-третьих, анальная лопасть, или пигидий, находящийся на самом заднем конце тела и представляющий собой неметамерный отдел, который образуется у личинок еще до метаморфоза. Впереди пигидия находится зона роста, где возникают новые сегменты. Анальная лопасть обычно снабжена двумя нитевидными анальными усиками, или уритами. У Aphrodita уриты отсутствуют, но на параподиях последних сегментов имеются удлиненные спинные усики, образующие бахрому вокруг ануса.
2.2.1 ГОЛОВНОЙ ОТДЕЛ
Передний, или головной, отдел тела состоит из простомиума (prostomium), иначе головной лопасти, и расположенного за ним первого (перистомиального) и второго (буккального) сегментов (рис. 3, 4). Первый сегмент сильно редуцирован со спины, и от него сохраняются лишь направленные вперед подиаль-ные выросты с перистомиальными, иначе тентакулярными усиками (cirri peri-stomiales, cirri tentaculares). Второй сегмент отличается от последующих туловищных сегментов в основном наличием очень длинных брюшных усиков, которые обозначаются как ротовые (cirri buccales), так как они расположены по бокам ротового отверстия и непосредственно участвуют в обнаружении и захвате пиши. Второй сегмент у всех чешуйчатых червей снабжен первой парой элитр, или спинных чешуек. На переднем крае II сегмента нередко имеется полукруглая складка, или вырост, заходящий сзади на спинную сторону головной лопасти (так называемая затылочная, или нухальная складка). Преобразования I и II сегментов в единый перистомиум путем их слияния у чешуйчатых червей не наблюдается. В этом отношении процессы цефализации у поли-ноид стоят на более низком уровне, чем у нереид, имеющих хорошо выраженный перистомиум в виде широкого ошейника, который представляет собой результат слияния трех передних ларвальных сегментов.
Простомиум у всех чешуйчатых червей хорошо развит и имеет крупные размеры. В процессе метаморфоза личинки он образуется непосредственно из верхнего полушария трохофоры (эписферы), т. е. имеет независимое от всех остальных сегментов происхождение. В соответствии с этим головная лопасть всегда располагается впереди ротового отверстия, находящегося на брюшной стороне червя. У чешуйчатых червей простомиум обычно вытянутой или округлой формы, иногда ширина его может быть больше длины. У переднего края головной лопасти находится 1—3 головных щупальца, или антенны (antennae): одно медиальное и два латеральных.
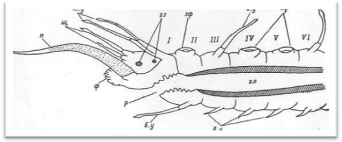
Рис. 3. Продольный разрез через передний отдел тела полиноиды.
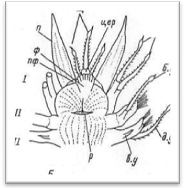
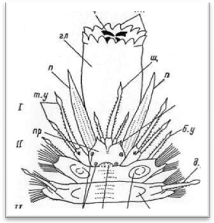
Рис. 4. Головной отдел полиноиды: вид сверху (А) и вид снизу (Б).
Головные щупальца веретенообразной формы, гладкие или с многочисленными мелкими папиллами. Щупальца большей частью имеют обособленные подставки, которые обозначаются как цератофоры, а сами щупальца — цератостили. Наличие цератофор обеспечивает большую подвижность самих щупалец. При медиальном щупальце иногда наблюдаются своеобразные ложковидные органы с ресничным эпителием — так называемые ктенидии (Laetmonice, Sthenelais и др.); назначение этих чувствительных органов точно не выяснено. Количество и характер расположения головных щупалец имеет большое систематическое значение: на этом основании в пределах семейства Polynoidae выделяются несколько подсемейств. У полиноид различают следующие типы головной лопасти (рис.5).
1.Лепидонотоидный тип – три головных щупальца; боковые дольки головной лопасти сильно вытянуты вперед и непосредственно (без базальных подставок) переходят в латеральные щупальца (терминальное положение латеральных щупалец); медиальное щупальце отходит от крупной базальной подставки (цератофора), расположенной в промежутке между боковыми выростами головной лопасти, и находится в одной плоскости с латеральными щупальцами.
2.Арктоноидный тип – три головных щупальца; латеральные щупальца имеют базальные подставки, отходящие от переднего края головной лопасти или несколько сдвинутые на брюшную сторону (субтерминальное положение);
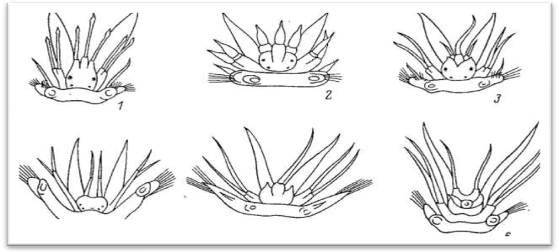
Рис.5 Основные типы головного отдела полиноид. 1 – лепидонотоидный, 2 – арктоноидный, 3 – хармотоидный, 4 – ифионоиднный, 5 – мацеллицефалоидный, 6 - адметеолоидный.
Подставка медиального щупальца лежит поверх латеральных. Арктоноидный тип является промежуточным между лепидонотоидным и хармотоидным.
3.Хармотоидный тип — три головных щупальца; латеральные щупальца с обособленными подставками - имеют вентральное положение, медиальное щупальце с крупной базальной подставкой сдвинуто на дорсальную сторону головной лопасти; дольки головной лопасти нередко с заостренными выростами, так называемыми лобными рогами.
4.Ифионоидный тип — имеются лишь латеральные щупальца, сидящие на боковых выростах головной лопасти; вместо медиального щупальца в задней части головной лопасти под нухальной складкой находится небольшая папилла.
5.Мацеллицефалоидный тип — имеется лишь одно хорошо выраженное медиальное щупальце, сильно сдвинутое на дорсальную сторону головной лопасти и сидящее на крупной базальной подставке (цератофоре); дольки головной лопасти округлые или с небольшими нитевидными придатками, которые представляют собой рудименты латеральных щупалец.
6.Адметеллоидный тип — три головных щупальца; латеральные щупальца лепидонотоидного типа, медиальное щупальце сдвинуто на спинную сторону головной лопасти, при нем имеются особые придатки — так называемые антеннальные чешуйки.
Головная лопасть у всех чешуйчатых червей со спины прикрывает первый (перистомиальный) сегмент, который из-за этого не имеет четких границ. При тентакулярных усиках первого сегмента, иногда обозначаемых как щупальцевидные усики, нередко имеются ацикула и пучок щетинок. Головные щупальца и тентакулярные усики внешне могут мало различаться, однако головные щупальца иннервируются от переднего отдела головного мозга, а перистомиальные усики — от коннективов ротового нервного кольца.
Помимо головных щупалец и тентакулярных усиков, у всех чешуйчатых червей имеются так называемые щупики, или пальпы (palpi), по своей массивности превосходящие все остальные головные придатки. Пальпы отходят с брюшной стороны головного отдела червя, по бокам от ротового отверстия, и внутрь их может заходить целомическая полость. Пальпы имеют очень небольшие подставки — пальпофоры, придающие им дополнительную подвижность. Вдоль наружной поверхности пальп обычно наблюдаются многочисленные продольные ряды мягких чувствительных папилл.
Все указанные здесь головные придатки являются осязательными органами и играют определенную роль при обнаружении пищи, но основным орудием захвата пищи служит выпячивающаяся наружу мускулистая глотка (pharynx), вооруженная нередко мощными хитиноидными челюстями.
На дорсальной стороне головной лопасти расположены глаза; они в большинстве случаев простого типа, без глазных стебельков (омматофоров). Глазные пятна темного (почти черного) цвета, округлой или овальной формы. У полиноид их две пары, причем первая (передняя) несколько крупнее задней, которая к тому же может быть частично прикрыта спинной складкой второго (буккаль-ного) сегмента. Первая пара глаз располагается по бокам головной лопасти в наиболее расширенной ее части или у переднего края, они могут частично переходить и на вентральную сторону головной лопасти (например, у Иаг-mothoe imbricata).
