Реферат: Биологическое время и его моделирование в квазихимическом пространстве
Методология построения теории времени естественных объектов, детально изложена [1, 2]. В данной работе рассмотрены компоненты этой теории на примере клеточной популяции.
1. Структура клеточной популяции. (O -компонент теории, object ).
Первая проблема состоит в выборе составных частей (элементов, таксонов, униструктур) объекта и определении их пространственных границ [3, 4].
Принципиальная трудность состоит в том, что строгий критерий членения объекта на части отсутствует. Выделение объектов природы – идеализация, связанная, главным образом, с особенностями информационных взаимодействий живых организмов со средой [5].
Объективным критерием членения объекта может служить принцип энергетической дифференцировки [6], который можно записать в виде:
Еj+1 << Еj ,
где Еj , Еj+1 — прочность связи между элементами уровня j и между элементами более высокого уровня j+1 (схема 1).
Для клеточной популяции порядок элементов с усложнением уровня (цифры - номера j уровней) можно представить в виде:
1 молекулы(t m )I 2 органеллы(t o ) I 3 клетки (t c ) I 4 популяция(t p ) I 5 экосистема(t e ) (1)
Каждый уровень j разбивается на подмножества элементов (схема 1).
Схема 1. Иерархия подсистем естественных объектов
В качестве высшего уровня в данной иерархии определена экосистема. В случае клеточной монокультуры экосистема состоит из двух частей: клеточной популяции и культуральной среды. Соответственно, в описание структуры объекта необходимо вводить описание структуры среды. В частности химическая структура среды включает набор питательных веществ (субстратов) и химических агентов:
|
Ms = (Ms1 , Ms2 , ... Mse ) Xs = (Xs1 , X2s , ... Xse ) |
(2) (3) |
где Ms , Xs - векторы (наборы) субстратов (Ms1 , Mse ) и химических агентов (Xs1 ,Xse ) для биологического вида S.
Многомерность структуры (1-3) создает слишком большие сложности не только для количественной, но и для качественной идентификации модели. Поэтому целесообразно исследовать агрегированные модели. В случае клеточной монокультуры весьма информативной оказывается структура (1) из трех уровней:
| (3) клетки I (4) популяция I (5) экосистема | (4) |
Описательная модель клеточного цикла [7], начиная с митоза Cm , может быть представлена в виде разветвленной цепи:
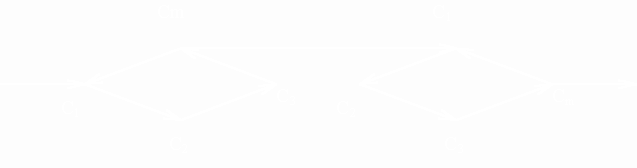 | (5a) |
или в более сжатом варианте:
| Cm ® f C1 ® C2 ® C3 ® Cm | (5b) |
Здесь C1 - молодая клетка непосредственно после деления, f - коэффициент размножения, который в общем случае может отличаться от 2. С2 , С 3 , Сm - фазы развития каждой из f клеток C1 .
Фазы C1 , C2 , C3 , Cm соответствуют стадиям клеточного цикла G1 , S, G2 , M (клетки четырех возрастов) и являются подмножествами 4-го уровня иерархии (4) - популяции. Популяция с наборами химических агентов (2) и (3) формирует 5-й уровень модели (4).
2. Пространство допустимых состояний популяции (S-компонент теории, states ).
Состояние системы отображается набором элементов (универсумов) всех типов (схема 1). В полной модели (2) состояние клеточной монокультуры задается числами всех типов мономеров, полимеров, органелл, клеток, химических и физических агентов среды. Размерность пространства состояний даже для культуры прокариотов составляет весьма большую величину.
Ограничения на достижимые состояния определяются условиями положительности числа компонентов и физической емкости среды.
В модели (4) состояние задается числами всех типов клеток, химических и физических агентов среды. Уменьшение размерности модели достигается путем аггрегирования разных элементов одного уровня по тому или иному признаку.
Например, в рассматриваемой ниже двухстадийной модели фазы C1 , C2 , C3 объединяются в одну по признаку их большой длительности и отсутствия деления на этих стадиях цикла. Разветвленный циклический граф, отображающий эту двухстадийную модель, имеет вид:
| (5c) |
3. Изменение состояний популяции, «предвремя» ( C-компонент теории, change).
--> ЧИТАТЬ ПОЛНОСТЬЮ <--