Реферат: Биологическое время и его моделирование в квазихимическом пространстве
С помощью различных приближений [11] система уравнений четвертого порядка может быть редуцирована до второго. Такая система достаточно информативна и позволяет качественно, а во многих случаях и количественно, описать развитие популяций различных видов.
Рассмотрим редуцированную модель (1) из двух стадий – роста и деления, дополненных стадией самоингибирования:
| C1 +M1 ® Cm | (p) | |
| Cm +M2 ® fC1 | (b) | |
| C1 ® Cd | (g) | |
| C1 +Cm ® Ca +C1 | (a) | (11) |
| C1 « EE | (w1 ) | |
| C1 +X1 « (C1 X11 ) | (d11 ) | |
| C1 +X2 « (C1 X12 ) | (d12 ) | |
| Cm +X1 « (Cm X21 ) | (d21 ) | |
| Cm +X2 « (Cm X22 ) | (d22 ) |
Здесь использованы те же обозначения, что и в системах (6) – (9) (индексы опущены): С1 – множество клеток разного возраста до митоза, Сm – митотические клетки; Ca – клетки в анабиозе; (Ck Xl ) – ингибированные клетки разных стадий; M1 , M2 –субстраты.
Следует отметить, что двухстадийный цикл (фазы S и M) наблюдается на ранних стадиях развития зародышей пойкилотермных животных [15, 16]. На этом основании в этот период в качестве единичного интервала времени можно использовать длительность tc клеточного цикла («детлаф»).
В предположении постоянства концентраций субстратов М1 , М2 кинетика цепного роста популяции, состоящей из особей С1 и Сm , описывается системой:
| dc1 /dt = – px c1 + f b cm + w1 | ( 12.1 ) |
| dcm /dt = p c1 – bx cm – a c1 cm | ( 12.2 ) |
Здесь c1 , cm – количества растущих и митотические клеток; a, b, p –коэффициенты автоингибирования, рождения и роста популяции в отсутствии ингибиторов. В коэффициенты р и b включены постоянные количества субстратов М1 и М2 . f - коэффициент размножения. Коэффициенты bx и px – функции количества ингибиторов x1 и x2 :
| px = p+d1 ; bx = b + d2 , где d1 = d11 x1 + d12 x2 ; d2 = d21 x1 + d22 x2 . (13) |
Система уравнений (12) представляет собой закон обобщенного движения двухстадийной популяции в пространстве состояний. Преобразованная в виде:
| dc1 = (-px c1 + f b cm + w1 ) dt, dcm =(p c1 - bx cm - a c1 cm ) dt, ( 12а ) |
система (12) определяет соотношение между интервалами биологического dci и физического dt времени.
В приближении квазистационарности для митотических клеток Сm система (12.1-12.2) сводится к одному уравнению:
| dc1 /dt=px c1 (K1 –c1 )/(K2 + c1 ) + w1 | (14) |
| Здесь K1 =c1 ``=(f b p - px bx )/(a px ); K2 =bx /a. | (15) |
Динамику численности популяции с1 (t) в общем случае нельзя выразить в виде явной функции от времени t. Поэтому используют обратную функцию t(c1 ), получаемую интегрированием (14) по с1 :
| t(c1 )=ln{(c1 /c0 )[(K1 -c0 )/(K1 -c1 )](1+n) }/(npx ), где n= K1 /K2 . (16) |
Уравнение (16) в явном виде отображает физическое время на множество состояний популяции.
6. Интерпретация модели (I-компонент теории, interpretation).
На рис.1 приведены экспериментальные точки и графики функции (16), описывающие дрожжевых клеток в присутствии солей хрома и никеля [13]. При расчете графиков брали значения a, b, р , f, определенные по экспериментальным данным [17]. В пределах точности измерений расчетные кривые согласуются с экспериментом при измененении численности примерно на шесть порядков.
Хорошее согласие теории с экспериментом получено и для других биологических объектов [8, 9]. Поэтому интересно провести верификацию квазихимической модели по характеристикам, связанным с проблемами биологического времени.
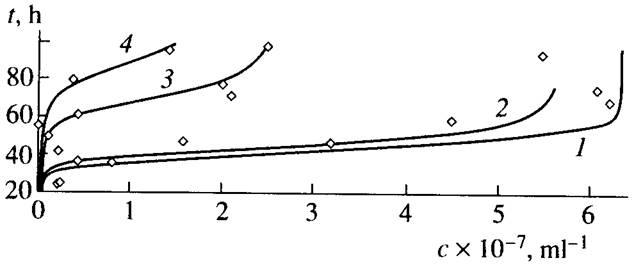
Рис.1. Экспериментальные точки и графики функции (16), описывающие рост пивных дрожжей при разных концентрациях (ммоль/л) солей хрома и никеля [13]: (1) c(Ni) = c(Cr) = 0.0, (2) c(Ni) = 0.5, (3) c(Cr) = 0.5, and (4) c(Ni) + c(Cr) = 0.5 + 0.5. Коэффициенты: a=1.25.10-7 мл /ч, b=0.8ч-1 , р=0.32 ч-1 , f =2.
Уравнения (12a) можно представить в виде:
| dc1 = Kc1 dt, dcm = Kcm dt, | ( 17) |
где величины Kc1 =(-px c1 +f b cm +w1 )и Kcm = (p c1 -bx cm -a c1 cm )представляют собой калибровочные коэффициенты для перехода от интервала физического времени dt к интервалам биологического времени dcj .
На основе (17) получают соотношение между конечными временными интервалами:
| D c1 = | ( 17а) |
Калибровочные соотношения (17) обладают следующими свойствами:
1. Коэффициенты Kc1 и Kcm зависят от кинетических констант, характеризующих внутри- и внесистемные взаимодействия. Это определяет специфику биологического времени данного объекта.
2. Коэффициенты Kc1 и Kcm зависят от наблюдаемого состояния объекта, то есть изменяются при движении по фазовой траектории.
3. Коэффициенты Kc1 и Kcm неодинаковы для однотипных элементов данного уровня иерархии. Это означает, что собственное время «течет» с разной скоростью не только на разных уровнях биологической системы, но и в различных элементах одного уровня.
Приращение суммарной массы dmp или численности dNp популяции определяют интервал биологического времени популяции в целом. Для двухстадийной популяции dNp = V(с1 +сm ), где V – объем системы. Связь между популяционным и физическим временем согласно (17) определяется соотношением:
| dNp = V( Kc1 + Kcm )dt . | ( 18) |
Через длительность клеточного цикла tc в физической шкале (в «детлафах») эта величина выразится в виде:
| dNpd = V( Kc1 + Kcm )dt / tc . | (19) |
Длительность клеточного цикла tc в физической шкале рассчитывают либо по экпериментальным значениям прироста массы или численности клеток, либо по экпериментальным значениям параметров b и p модели (12).
Приращение численности популяции D с12 =c2 -c1 в единице обьема наблюдается за время D t12 =t2 -t1 , согласно (16) равное:
| D t12 =ln{(c2 /c1 )[(K1 -c1 )/(K1 -c2 )](1+n) }/(npx ), | (20) |
где c1 ,c2 – численности в моменты t1 ,t2 .
Среднее число делений n12 каждой из c1 клеток за это время равно:
| n12 = log(c2 /c1 )/log2 | (21) |
Следователъно, t c можно оценить по формуле:
| tc = D t12 / n12 | (22) |