Реферат: Применение геометрического подхода в мембранной теории возбуждения
В начале 60-х годов натриевая теория ПД стала общепризнанной. Однако ее экспериментальная база была очень узкой — аксон кальмара. Поэтому в разных лабораториях мира начали изучать применимость этой теории к другим биологическим объектам. В большинстве случаев оказывалось, что самые разные возбудимые клетки — нервные волокна лягушки и кошки, мышечные волокна и др.— работают так же, как аксон кальмара.
Загадка миокардиальных клеток и «геометрический подход»
Одним из удивительных исключений оказались клетки сердца млекопитающих. Сначала и там все было в порядке. Швейцарский физиолог С. Вайдман изучал волокна Пуркинье проводящей системы сердца, которые передают сигнал от специальной структуры, лежащей на границе предсердия и желудочка, к мышечным клеткам желудочка. Эти волокна образованы «колонкой» клеток, соединенных высокопроницаемыми контактами. Вайдман показал, что в этих сердечных клетках быстрый подъем потенциала при возбуждении связан с открыванием натриевых каналов. Скорость нарастания ПД и его амплитуда зависели от содержания ионов натрия в наружном растворе, а сопротивление мембраны на пике ПД падало примерно в 100 раз по сравнению с состоянием покоя. Все было очень похоже на аксон кальмара.
Однако в случае клеток предсердия и желудочка ситуация оказалась совершенно иной и весьма неожиданной. И там амплитуда ПД зависела от концентрации ионов Ка+ в растворе, а ее значение составляло примерно 4-20 мВ; это означало, что проницаемость мембраны для 1Ча+ становилась примерно на порядок больше, чем для калия. Однако входное сопротивление ткани при возбуждении не менялось!
Этот факт прямо противоречил существующим взглядам. Для его объяснения придумывали разнообразные искусственные конструкции. Например, предполагали, что в сердечных клетках есть особые калиевые каналы, которые закрываются при деполяризации точно с той же скоростью, с какой натриевые открываются. В результате сопротивление не меняется. Но доказать существование таких быстрозакрывающихся каналов не удавалось. Неудачными оказывались и другие варианты объяснений.
В это время два биофизика С.А. Ковалев и Л. М. Чайлахян занимались еще одной задачей, казалось бы, не имеющей никакого отношения к сердцу. Они выясняли, как будет распространяться возбуждение по неоднородному нервному волокну. Вспомните, что в гл. 6 для простоты рассматривается бесконечный цилиндрический кабель, т. е. идеализированное однородное волокно. Но реальные нервные волокна могут расширяться и сужаться, практически все они ветвятся и обязательно где-то кончаются. Задача состояла в том, чтобы выяснить, что будет с нервным импульсом, когда он подойдет к области неоднородности — утолщению или мосту ветвления. Для таких случаев теории тогда не существовало.
Размышляя о ветвящихся волокнах, Ковалев и Чайлахян выдвинули совсем новую гипотезу для объяснения загадки сердечных клеток. Они рассуждали так. В проводящей системе сердца все происходит, как в аксоне кальмара, а в желудочке совсем иначе. Но проводящая система отличается от желудочка, прежде всего геометрической структурой: проводящая система образована волокнами, похожими на аксон дождевого червя,— они сходны с кабелем; желудочек, напротив, образован сложной сетью клеток, связанных высокопроницаемыми контактами, и никак не может считаться кабелем. Может быть, для объяснения необычного поведения желудочка не нужно искать какие-то необычные ионные каналы, но необходимо учесть эти геометрические различия?
Для ответа на этот вопрос надо было иметь для геометрически разных структур такую же теорию, как для однородного волокна.
С.А. Ковалев и Л.М. Чайлахян работали в Теоретическом отделе Института биофизики АН СССР, организованном в начале 60-х годов Г.М. Франком и И. М. Гельфандом. В нем собрались молодые математики, физики и биологи. Объединение разных знаний очень помогало в работе. Такое содружество и привело к тому, что в Теоретическом отделе было разработано обобщение кабельной теории, и в конце концов возник новый подход к возбудимым тканям, который для краткости назвали геометрическим.
В чем суть геометрического подхода? Свойства нервных и мышечных клеток во многом определяются свойствами их мембран. Эти свойства, в свою очередь, определяются тем, какие ионные каналы находятся в мембране. Однако, как оказалось, у природы есть и другой способ менять свойства клеток — менять их форму.
В биологии есть очень древняя проблема, которую называют проблемой формы и функции. Например, форма конечности у крота приспособлена для рытья земли, у летучей мыши — для полета, а у обезьяны — для хватания. С другой стороны, форма тела столь далеких по происхождению организмов, как древний ящер — ихтиозавр, рыба — тунец и млекопитающее — дельфин, весьма сходна, так как все они быстрые пловцы.
Геометрический подход — это фактически применение той же идеи к возбудимым тканям. Его основной принцип состоит в том, что свойства возбудимых клеток и тканей, а значит, и выполняемые ими функции во многом определяются их геометрической структурой: формой клеток, соотношением их размеров, взаимным расположением и связями.
Геометрический подход в электрофизиологии сейчас широко распространен и кажется вполне естественным: ведь поведение, например, нейрона существенно зависит от распределения потенциала на его мембране и от токов, которые текут в клетке и в окружающей ее среде. Но токи и напряжения, в свою очередь, зависят от распределения сопротивлений и емкостей, а это распределение определяется формой клетки. Эти простые соображения оказались очень продуктивными при применении к конкретным биологическим задачам. И дальше мы рассмотрим несколько примеров применения такого геометрического подхода к нервным клеткам, которые могут иметь разные размеры и форму, к нервным волокнам — аксонам и дендритам, — которые тоже не всегда сходны с проводом постоянного диаметра, и к клеточным системам.
О шаре и цилиндре
Сравним электрические свойства тела нервной клетки и нервного волокна, считая тело клетки шаром, а волокно — бесконечным цилиндрическим кабелем.
Сравнение электрических свойств шара и цилиндра, образованных одинаковыми мембранами, покажет нам, какую важную роль в определении этих свойств играет форма.
Электрические параметры, которыми характеризуются клетки и ткани, можно разделить на две группы. К первой группе относятся параметры, характеризующие вещество мембраны и протоплазмы: удельное сопротивление мембраны, которое обычно равно 1—10 кОм-см2 , ее удельная емкость, обычно равная 1 мкФ/см2 , и, наконец, удельное сопротивление протоплазмы, которое равно примерно 100 Ом-см. Эти параметры, естественно, не зависят от формы и размеров клетки.
Ко второй группе относятся параметры, которые называют системными. Они характеризуют клетку, волокно или систему связанных клеток в целом и зависят от размеров и формы. Один из основных системных параметров — это так называемое входное сопротивление, о котором нам уже приходилось говорить в гл. 6.
Входное сопротивление измеряется так. Один микро-электрод вводится в одиночную клетку, в волокно или в одну из клеток, электрически связанных с соседями, другой полюс при этом помещают в наружную среду и пропускают между ними ток /. Одновременно с этим второй микроэлектрод вводят в ту же одиночную клетку, волокно или клетку-соседа. Тогда при пропускании тока второй микроэлектрод будет регистрировать сдвиг МП А7, Второй микроэлектрод вводят возможно ближе к токовому электроду, поскольку, как мы знаем, в нервном волокне сдвиг потенциала будет убывать по мере удаления от токового электрода. Входным сопротивлением называют отношение сдвига потенциала к пропускаемому току!
![]()
Таким образом, входное сопротивление — это аналог обычного сопротивления участка цепи. Естественно, что Лвх клеток зависит от параметров первой группы и от формы и размеров клеток, а также их взаимного расположения. Например, как мы видели в гл. 6 ), входное сопротивление аксона выражается формулой
![]()
Здесь Лш и НI — удельные сопротивления мембраны и протоплазмы аксона, т. е. параметры первой группы; все остальное в формуле определяется геометрией волокна — предполагалось, что это цилиндр, у которого длина гораздо больше радиуса.
А теперь посмотрим, что будет, если из того же материала сделать другое «платье» — сферическую клетку радиуса а.
Можно показать, что у сферических клеток, даже у очень крупных, с диаметром порядка 1 мм, сопротивление протоплазмы составляет только 0,1% сопротивления мембраны. У клеток меньших размеров — а таковы все клетки в нервной системе позвоночных — сопротивление протоплазмы можно вообще не учитывать. Так что реально входное сопротивление сферической клетки зависит только от удельного сопротивления ее мембраны и от ее радиуса а:
![]()
Сравним теперь формулы и. У сферической клетки Явх прямо пропорционально Ям , поэтому если при возбуждении клетки удельное сопротивление мембраны уменьшится, например, в 36 раз, то и 7?вх уменьшится тоже в 36 раз. У волокна же Лвх пропорционально не Дм , а только![]() , поэтому если при возбуждении сопротивление мембраны уменьшится также в 36 разЛ то /?вх изменится только в 6 раз.
, поэтому если при возбуждении сопротивление мембраны уменьшится также в 36 разЛ то /?вх изменится только в 6 раз.
Сферическая клетка и цилиндрическое волокно существенно отличаются и по своим емкостным свойствам. Если на сферическую клетку подействует постоянный токг
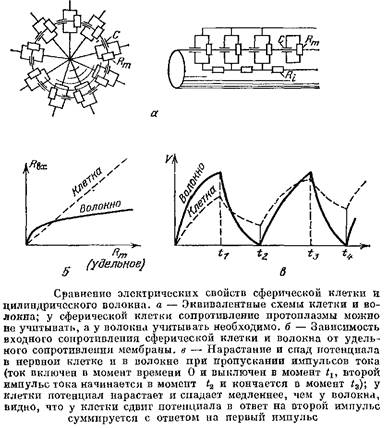
включенный в некоторый момент времени, то потенциал на ней будет нарастать по закону
![]()
--> ЧИТАТЬ ПОЛНОСТЬЮ <--