Реферат: Применение геометрического подхода в мембранной теории возбуждения
Заметим, что, поместив в область узла ветвления возбуждающие или тормозные синапсы, можно управлять процессом прохождения ПД через узел.
Зачем нейрону дендриты, а дендритам шипики
Многие нервные клетки похожи на кусты или деревья: их выходной отросток, аксон, — тонкий корешок этого дерева, все остальные многочисленные отростки — дендриты. Дендриты обычно отходят от тела клетки в виде толстых стволов, которые затем делятся на несколько более тонких ветвей, те, в свою очередь,— на еще более тонкие и т. д. Длина дендритов в десятки раз превышает диаметр нервных клеток, а толщина концевых веточек очень мала — может составлять доли микрометра. Вопрос о том, какую роль играют дендриты в работе нервных клеток, до сих пор окончательно не решен и, скорее всего, у разных нейронов их роль различна. В частности, у одних клеток мембрана дендритов невозбудима и может передавать сигналы только электротонически, как пассивный кабель, а у других — дендриты способны проводить ПД. Сейчас мы рассмотрим только те свойства дендритов, которые связаны с их геометрией.
Рассмотрим сначала те клетки, у которых дендриты невозбудимы. В этом случае «проблема дендритов» состоит в следующем. Синаптические окончания встречаются на самых разных частях дендритного дерева. Возьмем синапс, действующий на веточку, максимально удаленную от тела клетки. В этом случае условия для передачи электрического сигнала представляются весьма невыгодными. Действительно, в тонкой веточке велика константа затухания, а своим концом веточка «впадает» в более широкий участок дендрита, который «закорачивает» ее. В таких закороченных кабелях потенциал спадает особенно сильно, правда, в случае дендритов «закорачивание» является неполным и потенциал в конце веточки спадает не до нуля. В следующем отрезке дендрита условия для передачи сигнала тоже неблагоприятные, так как на его конце тоже находится более толстый дендритный ствол, и т. д. « В связи с этим возникло представление, что синапсы, расположенные на удаленных веточках, дают очень малый вклад в изменение потенциала тела клетки, в сотни раз меньший, чем такие же синапсы на теле клетки. Получается, что синапсы на конечных дендритных веточках бесполезны, что это «ошибка природы».
Один из вариантов решения «проблемы дендритов» состоит в том, что на тонких концевых веточках можно разместить много синапсов, тогда совместное действие этих синапсов будет заметно в теле клетки. Но для этого надо, чтобы все эти синапсы работали более или менее одновременно.
Все вышеприведенные рассуждения долгое время носили качественный характер. В 1965 г. в Теоретическом отделе Института биофизики АН СССР был разработан способ количественной оценки эффективности синапсов для нервных клеток любой формы и рассчитана эта эффективность для мотонейронов, пирамидных клеток коры и клеток мозжечка. Оказалось, что эффективность дендритных синапсов всего в 3—5 раз ниже, чем у синапсов, расположенных на теле нейрона. Чем это объясняется? Почему эффективность удаленных дендритных синапсов оказалось довольно велика? Чем меньше клетка, чем выше ее входное сопротивление, тем больший сдвиг потенциала создает синапс. У тоненьких дендритных веточек, удаленных от тела клетки, входное сопротивление оказалось большим, поэтому синапсы могут создавать в этих веточках сдвиги потенциала в десятки раз большие, чем в теле нейронов. И хотя при распространении к телу этот сдвиг потенциала действительно сильно затухает, его большая величина в значительной мере компенсирует затухание. Таким образом, дендритные синапсы оказались вовсе не ошибкой природы.
А теперь рассмотрим те нейроны, дендриты которых обладают возбудимой мембраной, способной к генерации ПД, У таких нейронов высокая эффективность синапса на тонкой веточке может привести к тому, что всего несколько синапсов доведут мембранный потенциал до порога и вызовут в этой веточке ПД, который начнет распространяться к телу клетки.
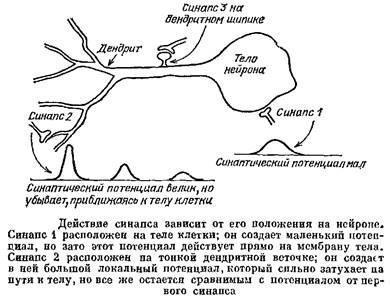
Его дальнейшая судьба зависит от свойств узлов ветвления, через которые ему надо пройти по пути к телу клетки, т. е. от геометрии дендрита. Клетка такого типа работает как сложная логическая схема. Пример такой клетки был приведен на рис, 45; эта клетка обнаруживает однонаправленные движения стимула. Клетки с более сложной формой дендритов могут работать как довольно хитрые вычислительные машины. «Такая система подобна системе голосования с большим числом участников, которые имеют неодинаковое число голосов... Окончательный результат, конечно, зависит от общего числа голосов, поданных «за» или «против», однако он в не меньшей степени зависит и от того, кто именно и вместе с кем из партнеров голосует»,— писали сотрудники Теоретического отдела Института биофизики АН СССР в 1966 г.
На дендритах многих нейронов имеются особые образования, так называемые шипики. Это структуры, похожие на грибы и состоящие из головки на тонкой ножке, которую чаще называют шейкой шипика. Шипик представляет собой выпячивание клеточной мембраны, а к его головке подходит терминаль от другого нейрона и образует на ней химический синапс.
Зачем нужны шипики — неизвестно. Число гипотез об их функциях огромно. Давайте посмотрим, что можно сказать о возможных функциях шипиков, исходя из геометрических соображений. При этом рассмотрим два ра-рианта: мембрана головки шипика невозбудима; мембрана головки шипика способна к генерации ПД.
Пусть шипик невозбудим. Его тоненькая шейка имеет высокое сопротивление. В результате в головке будет возникать большой постсинаптический потенциал, но его заметная часть будет теряться в шейке. Шипик будет работать как тоненькая дендритная веточка. Но зачем нужно такое устройство? Почему бы синапсу не располагаться прямо на дендрите?
Одним из способов работы тормозных синапсов является снижение входного сопротивления нейрона. Но ведь и возбуждающие синапсы тоже открывают ионные каналы и снижают входное сопротивление! Из-за этого возбуждающие синапсы тоже мешают друг другу. Особенно сильна такая помеха будет на тонких дендритах, у которых очень высоко входное сопротивление, так что активация нескольких синапсов вызовет заметное его снижение. Шипики должны существенно снижать взаимное влияние соседних синапсов, которые в этом случае отделены друг от друга шейками с высоким сопротивлением. Расчеты подтвердили, что хотя шипиковые синапсы каждый по отдельности менее эффективны, чем синапсы, расположенные прямо на дендрите, но при совместной работе эффект заметно выше.
Если же мембрана шипика возбудима, то он может работать как усилитель синаптической передачи. Из-за тонкости шейки входное сопротивление шипика очень велико и один синапс может вызвать в головке ПД, который пошлет в дендрит гораздо более сильный электрический ток, чем ток синапса. Интересно, что при таком режиме работы шипика должно существовать оптимальное сопротивление его шейки. Оно не должно быть слишком маленьким — тогда заметная часть синаптического тока будет утекать в дендритную веточку, сдвиг потенциала на мембране головки шипика не достигнет порогового значения и там не возникнет ПД. Но, с другой стороны, сопротивление шейки шипика не должно быть и слишком большим, иначе из головки шипика в дендрит будет течь слишком слабый ток и никакого усиления синаптического тока не получится. Недавно появились работы, показывающие, что геометрическая структура реальных шипиков близка к той,, которая по теоретическим расчетам является оптимальной.
До сих пор мы говорили о форме волокон и клеток или даже микроструктур клеток — шипиков. Посмотрим теперь на геометрию клеточных объединений.
Геометрическое выпрямление
Разберем сначала самый простой случай: пусть есть две клетки, соединенные между собой ЭС. Действие ЭС, в отличие от действия ХС, симметрично, И действительно, сам ЭС — это система тонких отверстий, которые совершенно одинаково пропускают ток в обе стороны. А вот действие такого синапса на связанные им клетки может сильно различаться, если эти клетки отличаются по величине, а значит, и по входному сопротивлению.
Пусть ЭС связывает две клетки — большую и маленькую. Пусть в какой-то из них возник сдвиг потенциала Д7Х . Этот сдвиг потенциала делится на две части — падение напряжения на сопротивлении ЭС и падение напряжения АУ2 на Д2 второй клетки, причем в соответствии с законом Ома потенциал распределяется пропорционально сопротивлениям участков. А отсюда сразу следуют важные выводы. Пусть,, например, Л2 первой клетки примерно в 9 раз меньше Лк , а -К2 второй клетки только в 4 раза меньше Лк - Если возбудилась первая клетка, то ее ПД, равный примерно 100 мВ, разделится в отношении 1:4, т. е. на второй клетке возникнет сдвиг потенциала около 20 мВ, и эта клетка возбудится, так как пороговое значение обычно равно примерно 15 мВ. Однако если такой же ІІД в 100 мВ возникнет во второй клетке, то он разделится в отношении 1: 9, тогда на долю первой клетки придется только одна десятая часть его, т, е. 10 мВ, что ниже порогового значения.
Получается, что ПД может передаваться через ЭС только в одном направлении. За счет разных размеров клеток возникает эффект, который называют геометрическим выпрямлением.
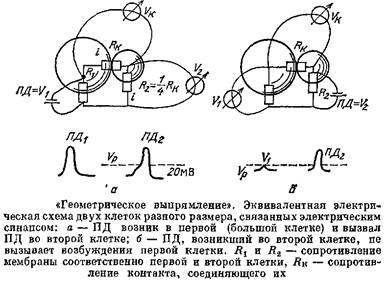
Но высокое и низкое Лвх зависит не только от размеров клеток. Пусть обе клетки однинаковы, но на одну из них подействовали химические синапсы, повысили проводимость ее мембраны, а это все равно, что сделали клетку большой. Таким образом, ХС могут регулировать работу ЭС и делать передачу через них односторонней.
Такая система регуляции существует, например, у некоторых моллюсков. Она управляет работой мышц их глотки. При необходимости схватить добычу моллюск делает резкое движение, все мышцы сокращаются одновременно» втягивая добычу. При этом все мотонейроны, управляющие мышцами, связаны ЭС, поэтому ПД в них возникает одновременно. При жевании те же мышцы должны работать неодновременно; это достигается тем, что под действием ХС Лвх мотонейронов снижается и ПД в одной клетке теперь уже не может возбудить другие.
Разгадка сердечной загадки
Необычное поведение миокарда: входное сопротивление в нем не меняется при возбуждении. Как оказалось, такое поведение миокарда объясняется не особенностями мембран сердечных клеток, а его собственной геометрией.
Клетки сердца соединены между собой многочисленными ЭС и образуют сложную сетку, так называемый синцитий.
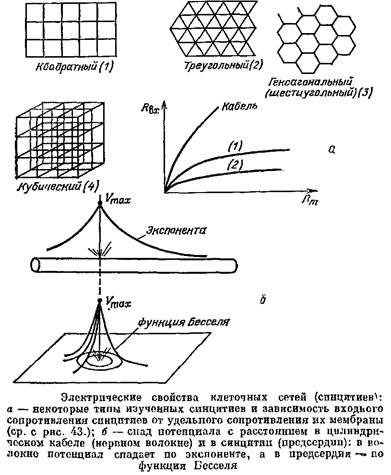
Для того чтобы выяснить электрические свойства синцитиев, надо было обобщить кабельную теорию и на этот случай. В качестве моделей миокарда были выбраны правильные сети. Отрезки кабеля между узлами моделировали миокардиальные клетки, а вся сеть, имеющая общий непрерывный сердечник и общую изоляцию, имитировала миокард. Основы математической теории таких синцитиев были созданы сотрудником Теоретического отдела Института биофизики АН СССР В.В. Смоляниновым, который впоследствии написал одну из первых книг о геометрическом подходе к возбудимым тканям.
Электрические свойства синцитиев оказались во многом необычными. Выяснилось, в частности, что Лвх синцитиев очень слабо зависит от сопротивления мембраны их клеток. Если даже*сопротивление мембраны упадет в 100 раз, то Явх изменится так мало, что это изменение не удастся обнаружить в эксперименте. Зато Явх синцитиев сильно зависит от сопротивления протоплазмы.
Зависимость Нвх геометрически разных структур от удельного сопротивления мембраны и удельного сопротивления протоплазмы.

Сравним теперь, как меняется мембранный потенциал в геометрически разных объектах по мере удаления от точечного источника тока. В сферической клетке сдвиг потенциала одинаков в любой точке ее мембраны — она эквипотенциальна. В цилиндрическом волокне потенциал спадает по экспоненте, а в синцитии потенциал спадает гораздо круче, чем по экспоненте; например, спад потенциала в таком почти плоском тонком синцитии, как предсердие лягушки, описывается функцией Бесселя.
Крутой спад потенциала в синцитиях позволяет, хотя fot на качественном уровне, объяснить, почему их /?вх так рлабо зависит от сопротивления мембраны. Пусть мы измеряемRBX ,пропуская ток между некоторой точкой синцития и наружной средой. Большая часть этого тока вытекает через область синцития, лежащую в некоторой окрестности точки пропускания тока; назовем эту область синцития эффективной. Пусть теперь сопротивление мембраны клеток синцития снизилось. От этого убывает константа длины волокон синцития. Потенциал спадает в каждом волокне круче, и площадь эффективной области синцития убывает. Практически эта площадь уменьшается примерно во столько же раз, во сколько раз снижается сопротивление мембраны. А так как сопротивление эффективной области тем меньше, чём меньше удельное сопротивление м